
Source: International Journal of Vaccine Theory Practice and Research
Stephanie Seneff1 and Greg Nigh2
1Computer Science and Artificial Intelligence Laboratory, MIT, Cambridge MA, 02139, USA, E-mail: seneff@csail.mit.edu
2Naturopathic Oncology, Immersion Health, Portland, OR 97214, USA
ABSTRACT
Operation Warp Speed brought to market in the United States two mRNA vaccines, produced by Pfizer and Moderna. Interim data suggested high efficacy for both of these vaccines, which helped legitimize Emergency Use Authorization (EUA) by the FDA.
However, the exceptionally rapid movement of these vaccines through controlled trials and into mass deployment raises multiple safety concerns.
In this review we first describe the technology underlying these vaccines in detail. We then review both components of and the intended biological response to these vaccines, including production of the spike protein itself, and their potential relationship to a wide range of both acute and long-term induced pathologies, such as blood disorders, neurodegenerative diseases and autoimmune diseases.
Among these potential induced pathologies, we discuss the relevance of prion-protein-related amino acid sequences within the spike protein. We also present a brief review of studies supporting the potential for spike protein “shedding”, transmission of the protein from a vaccinated to an unvaccinated person, resulting in symptoms induced in the latter.
We finish by addressing a common point of debate, namely, whether or not these vaccines could modify the DNA of those receiving the vaccination. While there are no studies demonstrating definitively that this is happening, we provide a plausible scenario, supported by previously established pathways for transformation and transport of genetic material, whereby injected mRNA could ultimately be incorporated into germ cell DNA for transgenerational transmission.
We conclude with our recommendations regarding surveillance that will help to clarify the long-term effects of these experimental drugs and allow us to better assess the true risk/benefit ratio of these novel technologies.
Keywords: antibody dependent enhancement, autoimmune diseases, gene editing, lipid nanoparticles, messenger RNA, prion diseases, reverse transcription, SARS-CoV-2 vaccines
Introduction
Unprecedented.
This word has defined so much about 2020 and the pandemic related to SARS-CoV-2.
In addition to an unprecedented disease and its global response, COVID-19 also initiated an unprecedented process of vaccine research, production, testing, and public distribution (Shaw, 2021).
The sense of urgency around combatting the virus led to the creation, in March 2020, of Operation Warp Speed (OWS), then-President Donald Trump’s program to bring a vaccine against COVID-19 to market as quickly as possible(Jacobs and Armstrong, 2020).
OWS established a few more unprecedented aspects of COVID-19.
First, it brought the US Department of Defense into direct collaboration with US health departments with respect to vaccine distribution (Bonsell, 2021).
Second, the National Institutes of Health (NIH) collaborated with the biotechnology company Moderna in bringing an unprecedented type of vaccine against infectious disease to market, one utilizing a technology based on messenger RNA (mRNA) (National Institutes of Health, 2020).
The confluence of these unprecedented events has rapidly brought to public awareness the promise and potential of mRNA vaccines as a new weapon against infectious diseases into the future.
At the same time, events without precedent are, by definition, without a history and context against which to fully assess risks, hoped-for benefits, safety, and long-term viability as a positive contribution to public health.
In this paper, we will be briefly reviewing one particular aspect of these unprecedented events, namely the development and deployment of mRNA vaccines against the targeted class of infectious diseases under the umbrella of “SARS-CoV-2.”
We believe many of the issues we raise here will be applicable to any future mRNA vaccine that might be produced against other infectious agents, or in applications related to cancer and genetic diseases, while others seem specifically relevant to mRNA vaccines currently being implemented against the subclass of coronaviruses.
While the promises of this technology have been widely heralded, the objectively assessed risks and safety concerns have received far less detailed attention.
It is our intention to review several highly concerning molecular aspects of infectious disease-related mRNA technology, and to correlate these with both documented and potential pathological effects.
Unprecedented
Many aspects of Covid-19 and subsequent vaccine development are unprecedented for a vaccine deployed for use in the general population. Some of these includes the following.
1. First to use PEG (polyethylene glycol) in an injection (see text)
2. First to use mRNAvaccine technology against an infectious agent
3. First time Moderna has brought any product to market
4. First to have public health officials telling those receiving the vaccination to expect an adverse reaction
5. First to be implemented publicly with nothing more than preliminary efficacy data (see text)
6. First vaccine to make no clear claims about reducing infections, transmissibility, or deaths
7. First coronavirus vaccine ever attempted in humans
8. First injection of genetically modified polynucleotides in the general populatio
Vaccine Development
Development of mRNA vaccines against infectious disease is unprecedented in many ways.
In a 2018 publication sponsored by the Bill and Melinda Gates Foundation, vaccines were divided into three categories: Simple, Complex, and Unprecedented (Young et al., 2018).
Simple and Complex vaccines represented standard and modified applications of existing vaccine technologies.
Unprecedented represents a category of vaccine against a disease for which there has never before been a suitable vaccine.
Vaccines against HIV and malaria are examples. As their analysis indicates, depicted in Figure 1, unprecedented vaccines are expected to take 12.5 years to develop.
Even more ominously, they have a 5% estimated chance of making it through Phase II trials (assessing efficacy) and, of that 5%, a 40% chance of making it through Phase III trials (assessing population benefit).
In other words, an unprecedented vaccine was predicted to have a 2% probability of success at the stage of a Phase III clinical trial.
As the authors bluntly put it, there is a “low probability of success, especially for unprecedented vaccines.” (Young et al., 2018)
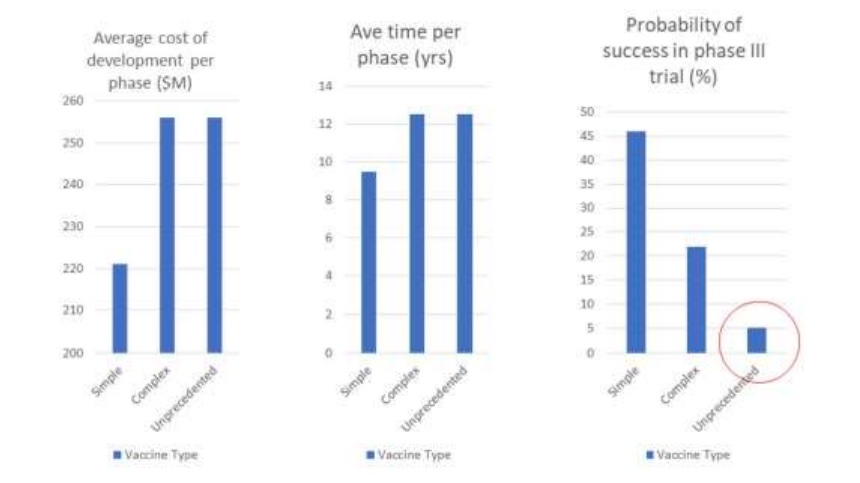
Figure 1. Launching innovative vaccines is costly and time-consuming, with a low probability of success, especially for unprecedented vaccines (adapted from Young et al, 2018).
With that in mind, two years later we have an unprecedented vaccine with reports of 90-95% efficacy (Baden et al. 2020).
In fact, these reports of efficacy are the primary motivation behind public support of vaccination adoption (U.S. Department of Health and Human Services, 2020).
This defies not only predictions, but also expectations. The British Medical Journal(BMJ) may be the only prominent conventional medical publication that has given a platform to voices calling attention to concerns around the efficacy of the COVID-19 vaccines.
There are indeed reasons to believe that estimations of efficacy are in need of re-evaluation. Peter Doshi, an associate editor of the BMJ, has published two important analyses (Doshi 2021a, 2021b) of the raw data released to the FDA by the vaccine makers, data that are the basis for the claim of high efficacy.
Unfortunately, these were published to the BMJ’s blog and not in its peer-reviewed content. Doshi, though, has published a study regarding vaccine efficacy and the questionable utility of vaccine trial endpoints in BMJ’s peer reviewed content (Doshi 2020).
A central aspect of Doshi’s critique of the preliminary efficacy data is the exclusion of over 3400 “suspected COVID-19 cases” that were not included in the interim analysis of the Pfizer vaccine data submitted to the FDA.
Further, a low-but-non-trivial percent of individuals in both Moderna and Pfizer trials were deemed to be SARS-CoV-1-positive at baseline despite prior infection being grounds for exclusion.
For these and other reasons the interim efficacy estimate of around 95% for both vaccines is suspect.
A more recent analysis looked specifically at the issue of relative vs. absolute risk reduction. While the high estimates of risk reduction are based upon relative risks, absolute risk reduction is a more appropriate metric for a member of the general public to determine whether a vaccination provides a meaningful risk reduction personally.
In that analysis, utilizing data supplied by the vaccine makers to the FDA, the Moderna vaccine at the time of interim analysis demonstrated an absolute risk reduction of 1.1% (p= 0.004), while the Pfizer vaccine absolute risk reduction was 0.7% (p<0.000) (Brown 2021).
Others have brought up important additional questions regarding COVID-19 vaccine development, questions with direct relevance to the mRNA vaccines reviewed here.
For example, Haidere, et. al. (2021) identify four “critical questions” related to development of these vaccines, questions that are germane to both their safety and their efficacy:
•Will Vaccines Stimulate the Immune Response?
•Will Vaccines Provide Sustainable Immune Endurance?
•How Will SARS-CoV-2 Mutate?
•Are We Prepared for Vaccine Backfires?
Lack of standard and extended preclinical and clinical trials of the two implemented mRNA vaccines leaves each of these questions to be answered over time.
It is now only through observation of pertinent physiological and epidemiological data generated by widescale delivery of the vaccines to the general public that these questions will be resolved.
And this is only possible if there is free access to unbiased reporting of outcomes –something that seems unlikely given the widespread censorship of vaccine-related information because of the perceived need to declare success at all cost.
The two mRNA vaccines that have made it through phase 3 trials and are now being delivered to the general population are the Moderna vaccine and the Pfizer-BioNTech vaccine.
The vaccines have much in common. Both are based on mRNA encoding the spike protein of the SARS-CoV-2 virus. Both demonstrated a relative efficacy rate of 94-95%.
Preliminary indications are that antibodies are still present after three months. Both recommend two doses spaced by three or four weeks, and recently there are reports of annual booster injections being necessary (Mahose, 2021).
Both are delivered through muscle injection, and both require deep-freeze storage to keep the RNA from breaking down.
This is because, unlike double-stranded DNA which is very stable, single-strand RNA products are apt to be damaged or rendered powerless at warm temperatures and must be kept extremely cold to retain their potential efficacy (Pushparajah et al., 2021).
It is claimed by the manufacturers that the Pfizer vaccine requires storage at -94 degrees Fahrenheit (-70 degrees Celsius), which makes it very challenging to transport it and keep it cold during the interim before it is finally administered.
The Moderna vaccine can be stored for 6 months at -4 degrees Fahrenheit (-20 degrees Celsius), and it can be stored safely in the refrigerator for 30 days following thawing (Zimmer et al., 2021).
Two other vaccines that are now being administered under emergency use are the Johnson & Johnson vaccine and the AstraZeneca vaccine.
Both are based on a vector DNA technology that is very different from the technology used in the mRNA vaccines. While these vaccines were also rushed to market with insufficient evaluation, they are not the subject of this paper so we will just describe briefly how they are developed.
These vaccines are based on a defective version of an adenovirus, a double-stranded DNA virus that causes the common cold.
The adenovirus has been genetically modified in two ways, such that it cannot replicate due to critical missing genes, and its genome has been augmented with the DNA code for the SARS-CoV-2 spike protein.
AstraZeneca’s production involves an immortalized human cell line called Human Embryonic Kidney (HEK) 293, which is grown in culture along with the defective viruses (Dicks et al., 2012). The HEK cell line was genetically modified back in the 1970s by augmenting its DNA with segments from an adenovirus that supply the missing genes needed for replication of the defective virus (Louis et al., 1997). Johnson & Johnson uses a similar technique based on a fetal retinal cell line.
Because the manufacture of these vaccines requires genetically modified human tumor cell lines, there is the potential for human DNA contamination as well as many other potential contaminants.
The media has generated a great deal of excitement about this revolutionary technology, but there are also concerns that we may not be realizing the complexity of the body’s potential for reactions to foreign mRNA and other ingredients in these vaccines that go far beyond the simple goal of tricking the body into producing antibodies to the spike protein.
In the remainder of this paper, we will first describe in more detail the technology behind mRNA vaccines. We devote several sections to specific aspects of the mRNA vaccines that concern us with regard to the potential for both predictable and unpredictable negative consequences.
We conclude with a plea to governments and the pharmaceutical industry to consider exercising greater caution in the current undertaking to vaccinate as many people as possible against SARS-CoV-2.
Technology of mRNA Vaccines
In the early phase of nucleotide-based gene therapy development, there was considerably more effort invested in gene delivery through DNA plasmids rather than through mRNA technology.
Two major obstacles for mRNA are its transient nature due to its susceptibility to breakdown by RNAses, as well as its known power to invoke a strong immune response, which interferes with its transcription into protein.
Plasmid DNA has been shown to persist in muscle up to six months, whereas mRNA almost certainly disappears much sooner.
For vaccine applications, it was originally thought that the immunogenic nature of RNA could work to an advantage, as the mRNA could double as an adjuvant for the vaccine, eliminating the arguments in favor of a toxic additive like aluminum.
However, the immune response results not only in an inflammatory response but also the rapid clearance of the RNA and suppression of transcription. So this idea turned out not to be practical.
There was an extensive period of time over which various ideas were explored to try to keep the mRNA from breaking down before it could produce protein. A major advance was the realization that substituting methyl-pseudouridine for all the uridine nucleotides would stabilize RNA against degradation, allowing it to survive long enough to produce adequate amounts of protein antigen needed for immunogenesis (Liu, 2019).
This form of mRNA delivered in the vaccine is never seen in nature, and therefore has the potential for unknown consequences.
The Pfizer-BioNTech and Moderna mRNA vaccines are based on very similar technologies, where a lipid nanoparticle encloses an RNA sequence coding for the full-length SARS-CoV-2 spike protein.
In the manufacturing process, the first step is to assemble a DNA molecule encoding the spike protein. This process has now been commoditized, so it’s relatively straightforward to obtain a DNA molecule from a specification of the sequence of nucleotides (Corbett et al., 2020).
Following a cell-free in vitro transcription from DNA, utilizing an enzymatic reaction catalyzed by RNA polymerase, the single-stranded RNA is stabilized through specific nucleoside modifications, and highly purified.
The company Moderna, in Cambridge, MA, is one of the developers of deployed mRNA vaccines for SARS-CoV-2. Moderna executives have a grand vision of extending the technology for many applications where the body can be directed to produce therapeutic proteins not just for antibody production but also to treat genetic diseases and cancer, among others.
They are developing a generic platform where DNA is the storage element, messenger RNA is the “software” and the proteins that the RNA codes for represent diverse application domains.
The vision is grandiose and the theoretical potential applications are vast (Moderna, 2020). The technology is impressive, but manipulation of the code of life could lead to completely unanticipated negative effects, potentially long term or even permanent.
SARS-CoV-2 is a member of the class of positive-strand RNA viruses, which means that they code directly for the proteins that the RNA encodes, rather than requiring a copy to an antisense strand prior to translation into protein.
The virus consists primarily of the single-strand RNA molecule packaged up inside a protein coat, consisting of the virus’s structural proteins, most notably the spike protein, which facilitates both viral binding to a receptor (in the case of SARS-CoV-2 this is the ACE2 receptor) and virus fusion with the host cell membrane. The SARS-CoV-2 spike protein is the primary target for neutralizing antibodies.
It is a class I fusion glycoprotein, and it is analogous to haemagglutinin produced by influenza viruses and the fusion glycoprotein produced by syncytial viruses, as well as gp160 produced by human immunodeficiency virus (HIV) (Corbett et al., 2020).
The mRNA vaccines are the culmination of years of research in exploring the possibility of using RNA encapsulated in a lipid particle as a messenger. The host cell’s existing biological machinery is co-opted to facilitate the natural production of protein from the mRNA.
The field has blossomed in part because of the ease with which specific oligonucleotide DNA sequences can be synthesized in the laboratory without the direct involvement of living organisms.
This technology has become commoditized and can be done at large-scale, with relatively low cost. Enzymatic conversion of DNA to RNA is also straightforward, and it is feasible to isolate essentially pure single-strand RNA from the reaction soup (Kosuri and Church, 2014).
1. Considerations in mRNA Selection and Modification
While the process is simple in principle, the manufacturers of mRNA vaccines do face some considerable technical challenges.
The first, as we’ve discussed, is that extracellular mRNA itself can induce an immune response which would result in its rapid clearance before it is even taken up by cells. So, the mRNA needs to be encased in a nanoparticle that will keep it hidden from the immune system.
The second issue is getting the cells to take up the nanoparticles. This can be solved in part by incorporating phospholipids into the nanoparticle to take advantage of natural pathways of lipid particle endocytosis.
The third problem is to activate the machinery that is involved in translating RNA into protein. In the case of SARS-CoV-2, the protein that is produced is the spike protein.
Following spike protein synthesis, antigen-presenting cells need to present the spike protein to T cells, which will ultimately produce protective memory antibodies (Moderna, 2020).
This step is not particularly straightforward, because the nanoparticles are mostly taken up by muscle cells, which, being immobile, are not necessarily equipped to launch an immune response.
As we will see, the likely scenario is that the spike protein is synthesized by muscle cells and then handed over to macrophages acting as antigen-presenting cells, which then launch the standard B-cell-based antibody-generating cascade response.
The mRNA that is enclosed in the vaccines undergoes several modification steps following its synthesis from a DNA template. Some of these steps involve preparing it to look exactly like a human mRNA sequence appropriately modified to support ribosomal translation into protein. Other modifications have the goal of protecting it from breakdown, so that sufficient protein can be produced to elicit an antibody response.
Unmodified mRNA induces an immune response that leads to high serum levels of interferon-α (IF-α), which is considered an undesirable response. However, researchers have found that replacing all of the uridines in the mRNA with N-methyl-pseudouridine enhances stability of the molecule while reducing its immunogenicity (Karikó et al. 2008; Corbett et al., 2020).
This step is part of the preparation of the mRNA in the vaccines, but, in addition, a 7-methylguanosine “cap” is added to the 5’ end of the molecule and a poly-adenine (poly-A) tail, consisting of 100 or more adenine nucleotides, is added to the 3’ end. The cap and tail are essential in maintaining the stability of the mRNA within the cytosol and promoting translation into protein (Schlake et al., 2012; Gallie, 1991).
Normally, the spike protein flips very easily from a pre-fusion configuration to a post-fusion configuration. The spike protein that is in these vaccines has been tweaked to encourage it to favor a stable configuration in its prefusion state, as this state provokes a stronger immune response (Jackson et al., 2020).
This was done via a “genetic mutation,” by replacing a critical two-residue segment with two proline residues at positions 986 and 987, at the top of the central helix of the S2 subunit (Wrapp et al., 2020).
Proline is a highly inflexible amino acid, so it interferes with the transition to the fusion state. This modification provides antibodies much better access to the critical site that supports fusion and subsequent cellular uptake.
But might this also mean that the genetically modified version of the spike protein produced by the human host cell following instructions from the vaccine mRNA lingers in the plasma membrane bound to ACE2 receptors because of impaired fusion capabilities? What might be the consequence of this? We don’t know.
Researchers in China published a report in Nature in August 2020 in which they presented data on several experimental mRNA vaccines where the mRNA coded for various fragments and proteins in the SARS-CoV-2 virus.
They tested three distinct vaccine formulations for their ability to induce an appropriate immune response in mice.
The three structural proteins, S (spike), M and E are minimal requirements to assemble a “virus-like particle” (VLP). Their hypothesis was that providing M and E as well as the S spike protein in the mRNA code would permit the assembly of VLPs that might elicit an improved immune response, because they more closely resemble the natural virus than S protein exposed on the surface of cells that have taken up only the S protein mRNA from the vaccine nanoparticles.
They were also hoping that critical fragments of the spike protein would be sufficient to induce immunity, rather than the entire spike protein, if viral-like particles could be produced through augmentation with M and E (Lu et al., 2020).
They confirmed experimentally that a vaccine containing the complete genes for all three proteins elicited a robust immune response that lasted for at least eight weeks following the second dose of the vaccine.
Its performance was far superior to that of a vaccine containing only the spike protein. Disappointingly, a vaccine that contained only critical components of the spike protein, augmented with the other two envelope proteins, elicited practically no response.
Moderna researchers have conducted similar studies with similar results. They concluded that the spike protein alone was clearly inferior to a formulation containing RNA encoding all three envelope proteins, and they hypothesized that this was due to the fact that all three proteins were needed to allow the cell to release intact virus-like particles, rather than to just post the spike protein in the plasma membrane.
The spike protein alone failed to initiate a T cell response in animal studies, whereas the formulation with all three proteins did (Corbett et al., 2020). The two emergency-approved vaccines only contain mRNA code for spike protein (without E or M), and there must have been a good reason for this decision, despite its observed poor performance.
It is possible that more sophisticated design of the lipid nanoparticle (see below) resulted in the ability to have the lipids serve as an adjuvant (similar to aluminum that is commonly added to traditional vaccines) while still protecting the RNA from degradation.
Another curious modification in the RNA code is that the developers have enriched the sequence in cytosines and guanines (Cs and Gs) at the expense of adenines and uracils (As and Us). They have been careful to replace only the third position in the codon in this way, and only when it does not alter the amino acid map (Hubert, 2020).
It has been demonstrated experimentally that GC-rich mRNA sequences are expressed (translated into protein) up to 100-fold more efficiently than GC-poor sequences (Kudla et al., 2006).
So this appears to be another modification to further assure synthesis of abundant copies of the spike protein. We do not know the unintended consequences of this maneuver. Intracellular pathogens, including viruses, tend to have low GC content compared to the host cell’s genome (Rocha and Danchin, 2020).
So, this modification may have been motivated in part by the desire to enhance the effectiveness of the deception that the protein is a human protein.
All of these various modifications to the RNA are designed to make it resist breakdown, appear more like a human messenger RNA protein-coding sequence, and efficiently translate into antigenic protein.
2. Lipid Nanoparticle Construction
Lipid nanoparticles (LNPs), also known as liposomes, can encapsulate RNA molecules, protecting them from enzymatic degradation by ribonucleases, and thus they form an essential ingredient of a successful delivery method (Wadhwa et al., 2020; Xu et al., 2020).
These artificial constructs closely resemble exosomes. Exosomes are extracellular vesicles secreted by cells and taken up by their neighbors, and they also often embed DNA or RNA. Thus, these nanoparticles can take advantage of natural endocytosis processes that normally internalize extracellular exosomes into endosomes.
As the endosome acidifies to become a lysosome, the mRNA is released into the cytoplasm, and this is where translation into protein takes place. Liposomes have actually been found to be more successful at enhancing antigen presentation and maturation of dendritic cells, when compared to fusion proteins that encapsulate virus-based vaccines (Norling et al., 2019).
The lipid nanoparticles (LNPs) in these vaccines are composed of ionizable cationic lipids, phospholipids, cholesterol and polyethylene glycol (PEG). Together, this mixture assembles into a stable lipid bilayer around the mRNA molecule.
The phospholipids in these experimental vaccines consist of a phosphatidylcholine headgroup connected to two saturated alkyl tails through a glycerol linker. The lipid used in these vaccines, named 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), has 18 repeat carbon units. The relatively long chain tends to form a gel phase rather than a fluid phase.
Molecules with shorter chains (such as a 12-carbon chain) tend to stay in a fluid phase instead. Gel phase liposomes utilizing DSPC have been found to have superior performance in protecting RNA from degradation because the longer alkyl chains are much more constrained in their movements within the lipid domain.
They also appear to be more efficient as an adjuvant, increasing the release of the cytockines tumor necrosis factor-α (TNF-α), interleukin (IL)-6 and IL-1β from exposed cells (Norling et al., 2019).
However, their ability to induce an inflammatory response may be the cause of the many symptoms people are experiencing, such as pain, swelling, fever and sleepiness.
A study published in bioRxiv verified experimentally that these ionizable cationic lipids in lipid nanoparticles induce a strong inflammatory response in mice (Ndeupen et al., 2021).
The current mRNA vaccines are delivered through intramuscular injection. Muscles contain a large network of blood vessels where immune cells can be recruited to the injection site (Zeng et al., 2020). Muscle cells generally can enhance an immune reaction once immune cells infiltrate, in response to an adjuvant (Marino et al., 2011). Careful analysis of the response to an mRNA vaccine, administered to mice, revealed that antigen is expressed initially within muscle cells and then transferred to antigen-presenting cells, suggesting “cross-priming” as the primary path for initiating a CD8 Tcell response (Lazzaro et al., 2015).
One can speculate that muscle cells make use of an immune response that is normally used to deal with misfolded human proteins. Such proteins induce upregulation of major histocompatibility complex (MHC) class II proteins, which then bind to the misfolded proteins and transport them intact to the plasma membrane (Jiang et al., 2013).
The MHC-bound surface protein then induces an inflammatory response and subsequent infiltration of antigen-presenting cells (e.g., dendritic cells and macrophages) into the muscle tissue, which then take up the displayed proteins and carry them into the lymph system to present them to T-cells.
These T-cells can then finally launch the cascade that ultimately produces memory antibodies specific to the protein. Muscle cells do express MHC class II proteins (Cifuentes-Diaz et al., 1992).
As contrasted with class I, class II MHC proteins specialize in transporting intact proteins to the surface as opposed to small peptide sequences derived from the partial breakdown of the proteins (Jiang et al., 2013).
An in vitro study on non-human primates demonstrated that radiolabeled mRNA moved from the injection site into the draining lymph node and remained there for at least 28 hours.
Antigen-presenting cells (APCs) in both the muscle tissue as well as the draining lymph nodes were shown to contain radiolabeled mRNA (Lindsay et al., 2019).
Classical APCs include dendritic cells, macrophages, Langerhans cells (in the skin) and B cells. Many of the side effects associated with these vaccines involve pain and inflammation at the injection site, as would be expected given the rapid infiltration of immune cells.
Lymphadenopathy is an inflammatory state in the lymph system associated with swollen lymph nodes. Swollen lymph nodes in the arm pit (axillary lymphadenopathy) is a feature of metastatic breast cancer.
A paper published in 2021 described four cases of women who developed axillary lymphadenopathy following a SARS-CoV-2 vaccine (Mehta et al., 2021). The authors urged caution in misinterpreting this condition as an indicator requiring biopsy follow-up for possible breast cancer.
This symptom corroborates tracer studies showing that the mRNA vaccine is predominantly taken up by APCs that then presumably synthesize the antigen (spike protein) from the mRNA and migrate into the lymph system, displaying spike protein on their membranes.
A list of the most common adverse effects reportedby the FDA that were experienced during the Pfizer-BioNTech clinical trials include “injection site pain, fatigue, headache, muscle pain, chills, joint pain, fever, injection site swelling, injection site redness, nausea, malaise, and lymphadenopathy.” (US Food and Drug Administration, 2021).
We turn now to individual molecular and organ system concerns that arise with these mRNA vaccines.
Adjuvants, Polyethylene Glycol, and Anaphylaxis
Adjuvants are vaccine additives intended to “elicit distinctive immunological profiles with regard to the direction, duration, and strength of immune responses” from the vaccines to which they are added (Liang et al., 2020).
Alum or other aluminum compounds are most commonly utilized in traditional vaccines, and they elicit a wide range of systemic immune activation pathways as well as stromal cell activation at the site of the injection (Lambrecht et al., 2009; Danielsson & Eriksson, 2021).
An aluminum-based adjuvant was determined not to be optimal for a coronavirus vaccine, so other solutions were sought (Liang et. al., 2020).
A solution presented itself in the form of the widely used pharmaceutical ingredient polyethylene glycol, or PEG. A limiting factor in the use of nucleic-acid-based vaccines is the tendency for the nucleic acids to be quickly degraded by nuclease enzymes (Ho et al., 2021).
Regarding the RNAse enzymes targeting injected mRNA, these enzymes are widely distributed both intracellularly (primarily within the lysosomes) (Fujiwara et al., 2017) and extracellularly (Lu et al., 2018). To overcome this limitation, both mRNA vaccines currently deployed against COVID-19 utilize lipid-based nanoparticles as delivery vehicles.
The mRNA cargo is placed inside a shell composed of synthetic lipids and cholesterol, along with PEG to stabilize the mRNA molecule against degradation. The vaccine produced by Pfizer/BioNTech creates nanoparticles from 2-[(polyethylene glycol)-2000]-N,N-ditetradecylacetamide, or ALC-0159, commonly abbreviated simply as PEG (World Health Organization, 2021, January 14).
The Moderna vaccine contains another PEG variant, SM-102, 1,2-dimyristoyl-rac-glycero-3-methoxypolyethylene glycol2000 (World Health Organization, 2021, January 19).
For convenience we will abbreviate both PEG-modified lipids as PEG, and refer to the vaccines as PEGylated according to standard nomenclature.
The lipid shell plays a triple role. First, it protects the genetic material from degradation prior to cellular uptake. Second, the lipid shell, which also contains cholesterol, facilitates cellular uptake through fusion with the lipid membrane of the cell and subsequent endocytosis of the lipid particle, invoking naturally existing processes. And finally, it acts as an adjuvant (Ho et al., 2021).
It is in this latter role as immune stimulant that most concerns have been raised regarding the widespread use of PEG in an injection therapy.
In an article published in May 2019, prior to large clinical trials involving these PEGylated vaccines, Mohamed et. al. (2019) described a number of concerning findings regarding PEG and the immunological activation it had been shown to produce, which includes humoral, cell-mediated, and complement-based activation.
They note that, paradoxically, large injection doses of PEG cause no apparent allergic reaction. Small doses, though, can lead to dramatic pathological immune activation. l
Vaccines employing PEGylation utilize micromolar amounts of these lipids, constituting this potentially immunogenic low-dose exposure.
In animal studies it has been shown that complement activation is responsible for both anaphylaxis and cardiovascular collapse, and injected PEG activates multiple complement pathways in humans as well.
The authors of one study conclude by noting that “This cascade of secondary mediators substantially amplifies effector immune responses and may induce anaphylaxis in sensitive individuals. Indeed, recent studies in pigs have demonstrated that systemic complement activation (e.g., induced following intravenous injection of PEGylated liposomes) can underlie cardiac anaphylaxis where C5a played a causal role.” (Hamad et al., 2008)
It is also important to note that anaphylactoid shock in pigs occurred not with first injected exposure, but following second injected exposure (Kozma et al., 2019). The presence of antibodies against PEG is widespread in the population (Zhou et al., 2020). Yang and Lai (2015) found that around 42% of blood samples surveyed contained anti-PEG antibodies, and they warn that these could have important consequencesfor any PEG-based therapeutics introduced.
Hong et. al. (2020) found anti-PEG antibodies with a prevalence up to 72% in populations with no prior exposure to PEG-based medical therapy.
Lila et. al. (2018) note that the “existence of such anti-PEG antibodies has been intimately correlated with an impairment of therapeutic efficacy in tandem with the development of severe adverse effects in several clinical settings employing PEGylated-based therapeutics.”
Anaphylaxis to vaccines has previously been assumedto be rare based on the frequency of such events reported to VAERS, a database established by the Centers for Disease Control and Prevention in 1990 for reporting of adverse events related to vaccines (Centers for Disease Control and Prevention, 1990; Su et al., 2019).
While rare, anaphylaxis can be life-threatening, so it is important to monitor for the possibility in the short period following vaccination (McNeil et al., 2016).Sellaturay et. al., after reviewing 5 cases of anaphylaxis they link to PEG exposure, one near-fatal and involving cardiac arrest, write, “PEG is a high-risk ’hidden’ allergen, usually unsuspected and can cause frequent allergic reactions due to inadvertent re-exposure.
Allergy investigation carries the risk of anaphylaxis and should be undertaken only in specialist drug allergy centres.” (Sellaturay et al., 2020). In fact it has already been demonstrated that pre-existing antibodies to PEG are linked to more common and more severe reactions upon re-exposure (Ganson et al., 2016).
Is anaphylaxis upon exposure to PEG happening with a frequency relevant to public health? Numerous studies have now documented the phenomenon (Lee et al., 2015; Povsic et al., 2016; Wylon et al., 2016).
Anaphylactic reactions to the mRNA vaccines are widely reported in the media (Kelso, 2021) and, as noted above, have been frequently reported in the VAERS database (690 reports of anaphylaxis following SARS-CoV-2 vaccines up to January 29, 2021).
There are also some initial case studies published in the peer-reviewed literature (Garvey & Nasser, 2020; CDC COVID-19 Response Team, 2021, January 15). Anaphylaxis reactions to vaccines prior to these COVID-19 vaccines were generally reported at rates less than 2 cases per million vaccinations (McNeil et al., 2016), while the current rate with the COVID-19 vaccinations was reported by the CDC to be more than 11 cases per million (CDC COVID-19 Response Team, 2021, January 29).
However, a published prospective study on 64,900 medical employees, where their reactions to their first mRNA vaccination were carefully monitored, found that 2.1% of the subjects reported acute allergic reactions.
A more extreme reaction involving anaphylaxis occurred at a rate of 247 per million vaccinations (Blumenthal et al., 2021). This is more than 21 times as many as were initially reported by the CDC.
The second injection exposure is likely to cause even larger numbers of anaphylactic reactions.
mRNA Vaccines, Spike Proteins, and Antibody-Dependent Enhancement (ADE)
ADE is an immunological phenomenon first described in 1964 (Hawkes et al., 1964). In that publication Hawkes described a set of experiments in which cultures of flavivirus were incubated with avian sera containing high titers of antibodies against those viruses.
The unexpected finding was that, with increasingly high dilutions of the antibody-containing sera, cell infectivity was enhanced. Lack of an explanation for how this could happen is likely responsible for its being largely ignored for almost 20 years (Morens et al., 1994).
Multiple pathways have been proposed through which antibodies both directly and indirectly participate in the neutralization of infections (Lu et al., 2018b).
ADE is a special case of what can happen when low, non-neutralizing levels of either specific or cross-reactive antibodies against a virus are present at the time of infection. These antibodies might be present due to prior exposure to the virus, exposure to a related virus, or due to prior vaccination against the virus.
Upon reinfection, antibodies in insufficient numbers to neutralize the virus nevertheless bind to the virus. These antibodies then dock at the Fc receptor on cell surfaces, facilitating viral entry into the cell and subsequently enhancing the infectivity of the virus (Wan et. al.,2020).
ADE is believed to underlie the more severe dengue fever often observed in those with previous exposure (Beltramello et al., 2010), and might also play a role in more severe disease among those previously vaccinated against the disease (Shukla et al., 2020).
ADE is also believed to play a role in Ebola (Takada et al., 2003), zika virus infection (Bardina et al., 2017), and other flavivirus infections (Campos et al., 2020).
In an extended correspondence published in Nature Biotechnology, Eroshenko et. al. offer a comprehensive review of evidence suggesting that ADE could become manifest with any vaccinations employed against SARS-CoV-2.
Importantly, they note that ADE has been observed with coronavirus vaccines tested in both in Vitro and in Vivo models (Eroshenko et al., 2020). Others have warned about the same possibility with SARS-CoV-2 vaccines.
A theory for how ADE might occur in the case of a SARS-CoV-2 vaccine suggests that non-neutralizing antibodies form immune complexes with viral antigens to provoke excessive secretion of pro-inflammatory cytokines, and, in the extreme case, a cytokine storm causing widespread local tissue damage (Lee et al., 2020).
One extensive review of ADE potentially associated with SARS-CoV-2 vaccines noted, “At present, there are no known clinical findings, immunological assays or biomarkers that can differentiate any severe viral infection from immune-enhanced disease, whether by measuring antibodies, T cells or intrinsic host responses” (Arvin et al. 2020; Liu et al.,2019). We will return to this point again below.
Preexisting immunoglobulin G (IgG) antibodies, induced by prior vaccination, contribute to severe pulmonary damage by SARS-CoV in macaques (Liu et al., 2019).
Peron and Nakaya (2020) provide evidence suggesting that the much more diverse range of prior exposures to coronaviruses experienced by the elderly might predispose them to ADE upon exposure to SARS-CoV-2.
A concerning pre-print article reported that plasma from 76% of patients who had recovered from severe COVID-19 disease, when added to cultures of SARS-CoV-2 and susceptible cells, exhibited enhanced ability for SARS-CoV-2 viral infection of Raji cells (Wu et al., 2020).
The authors note that “the antibody titers [against the spike protein] were higher in elderly patients of COVID-19, and stronger antibody response was associated with delayed viral clearance and increased disease severity in patients. Hence it is reasonable to speculate that S protein-specific antibodies may contribute to disease severity during SARS-CoV-2 infection.” (Wu et al., 2020)
It has been reported that all three US vaccine manufacturers –Moderna, Pfizer, and Johnson & Johnson –are working to develop booster shots (Zaman 2021).With tens of millions of young adults and even children now with vaccine-induced coronavirus spike protein antibodies, there exists the possibility of triggering ADE related to either future SARS-CoV-2 infection or booster injection among this younger population. Time will tell.
The mRNA vaccines ultimately deliver the highly antigenic spike protein to antigen-presenting cells. As such, monoclonal antibodies against the spike protein are the expected outcome of the currently deployed mRNA vaccines.
Human spike protein monoclonal antibodies were found to produce high levels of cross-reactive antibodies against endogenous human proteins (Vojdani et. al., 2021; reviewed in more detail below).
Given evidence only partially reviewed here, there is sufficient reason to suspect that antibodies to the spike protein will contribute to ADE provoked by prior SARS-CoV-2 infection or vaccination, which may manifest as either acute or chronic autoimmune and inflammatory conditions.
We have noted above that it is not possible to distinguish an ADE manifestation of disease from a true, non-ADE viral infection. In this light, it is important to recognize that, when diseases and deaths occur shortly after vaccination with an mRNA vaccine, it can never be definitively determined, even with a full investigation, that the vaccine reaction was not a proximal cause.
Pathogenic Priming, Multisystem Inflammatory Disease, and Autoimmunity
Pathogenic priming is a concept that is similar in outcome to ADE, but different in the underlying mechanism. We discuss it here as a unique mechanism through which the mRNA vaccines could provoke associated pathologies.
In April 2020 an important paper was published regarding the potential for self-reactive antibodies to be generated following exposure to the spike protein and other antigenic epitopes spread over the length of SARS-CoV-2.
Lyons-Weiler (2020) coined the phrase “pathogen priming” because he believed the more commonly used “immune enhancement” fails to capture the severity of the condition and its consequences.
In his in silico analysis, Lyons-Weiler compared all antigenic SARS-CoV-2 protein epitopes flagged in the SVMTriP database (http://sysbio.unl.edu/SVMTriP/) and searched the p-BLAST database (https://blast.ncbi.nlm.nih.gov/Blast.cgi) for homology between those epitopes and endogenous human proteins.
Of the 37 SARS-CoV-2 proteins analyzed, 29 had antigenic regions. All but one of these 29 had homology with human proteins (putative self-antigens)and were predicted to be autoreactogenic.
The largest number of homologies were associated with the spike (S) protein and the NS3 protein, both having 6 homologous human proteins.
Functional analysis of the endogenous human proteins homologous with viral proteins found that over 1/3 of them are associated with the adaptive immune system.
The author speculates that prior virus exposure or prior vaccination, either of which could initiate antibody production that targets these endogenous proteins, may be playing a role in the development of more severe disease in the elderly in particular.
In this case, the pre-existing antibodies act to suppress the adaptive immune system and lead to more severe disease.
Another group (Ehrenfeld et. al., 2020), in a paper predominantly about the wide range of autoimmune diseases found in association with a prior SARS-CoV-2 infection, also investigated how the spike protein could trigger such a range of diseases.
They report, in Table 1 of that reference, strings of heptapeptides within the human proteome that overlap with the spike protein generated by SARS-CoV-2. They identified 26 heptapeptides found in humans and in the spike protein.
It is interesting to note that 2 of the 26 overlapping heptapeptides were found to be sequential, a strikingly long string of identical peptides to be found in common between endogenous human proteins and the spike protein.
Commenting on the overlapping peptides they had discovered and the potential for this to drive many types of autoimmunity simultaneously, they comment, “The clinical scenario that emerges is upsetting.” Indeed, it is.
In May of 2020, another important paper in this regard was published by Vojdani and Kharrazian (2020). The authors used both mouse and rabbit monoclonal antibodies against the 2003 SARS spike protein to test for reactivity against not only the spike protein of SARS-CoV-2, but also against several endogenous human proteins.
They discovered that there was a high level of binding not only with the SARS-CoV-2 spike protein, but against a wide range of endogenous proteins. “[W]e found that the strongest reactions were with transglutaminase 3 (tTG3), transglutaminase 2 (tTG2), ENA, myelin basic protein (MBP), mitochondria, nuclear antigen (NA), α-myosin, thyroid peroxidase (TPO), collagen, claudin 5+6, and S100B.” (Vojdani and Kharrazian, 2020).
These important findings need to be emphasized.
Antibodies with a high binding affinity to SARS-CoV-2 spike and other proteins also have a high binding affinity with tTG (associated with Celiac Disease), TPO (Hashimoto’s thyroiditis), myelin basic protein (multiple sclerosis), and several endogenous proteins.
Unlike the autoimmune process associated with pathogen priming, these autoimmune diseases typically take years to manifest symptomatically.
The autoantibodies generated by the spike protein predicted by Lyons-Weiler (2020) and described above were confirmed with an in vitro study published more recently. In this follow-on paper, Vojdani et. al., (2021) looked again at the issue of cross-reactivity of antibodies, this time using human monoclonal antibodies (mAbs) against the SARS-CoV-2 spike protein rather than mouse and rabbit mAbs.
Their results confirmed and extended their prior findings. “At a cutoff of 0.32 OD [optical density], SARS-CoV-2 membrane protein antibody reacted with 18 out of the 55 tested antigens.” These 18 endogenous antigens encompass reactivity to tissue in liver, mitochondria, the nervous and digestive system, the pancreas, and elsewhere in the body.
In a report on multisystem inflammatory syndrome in children (MIS-C), Carter et. al. (2020) studied 23 cases. Seventeen of 23 (68%) patients had serological evidence of prior SARS-CoV-2 infection.
Of the three antibodies assessed in the patient population (nucleocapsid, RBD, and spike), IgG spike protein antibody optical density (which quantifies antibody concentrations against a standardized curve (Wikipedia, 2021)), was highest (see Figure 1d in Carter et al., 2020).
MIS-C is now commonly speculated to be an example of immune priming by prior exposure to SARS-CoV-2 or to other coronaviruses. Buonsenso et. al. (2020) reviewed multiple immunologic similarities between MIS-C and disease related to prior β-hemolytic Group A streptococcal infection (GAS).
The authors write, “We can speculate that children’s multiple exposition to SARS-CoV-2 with parents with COVID-19 can work as a priming of the immune system, as happens with GAS infection and, in genetically predisposed children, lead to [MIS-C] development.
Anotherhypothesis is that previous infections with other coronaviruses, much more frequent in the pediatric population, may have primed the child immune system to SARS-CoV-2 virus.”
In June 2019 Galeotti and Bayry (2020) reviewed the occurrence of both autoimmune and inflammatory diseases in patients with COVID-19. They focus their analysis on MIS-C. After reviewing several previously published reports of a temporal link between COVID-19 and onset of MIS-C and describing a number of possible mechanistic connections between the two, the authors noted that no causal link had been established.
In a somewhat prescient recommendation, they wrote, “A fine analysis of homology between various antigens of SARS-CoV-2 and self-antigens, by use of in silico approaches and validation in experimental models, should be considered in order to confirm this hypothesis.”
It is precisely this type of in silico analysis carried out by Lyons-Weiler (2020) and by Ehrenfeld et. al. (2020) described in the opening paragraphs of this section which found the tight homology between viral antigens and self-antigens. While this may not definitively confirm the causal link hypothesized by Galeotti and Bayry, it is strong supporting evidence.
Autoimmunity is becoming much more widely recognized as a sequela of COVID-19.
There are multiple reports of previously healthy individuals who developed diseases such as idiopathic thrombocytopenic purpura, Guillain-Barré syndrome and autoimmune haemolytic anaemia (Galeotti and Bayry, 2020).
There are three independent case reports of systemic lupus erythemosus (SLE) with cutaneous manifestations following symptomatic COVID-19. In one case a 39-year-old male had SLE onset two months following outpatient treatment for COVID-19 (Zamani et.al., 2021).
Another striking case of rapidly progressing and fatal SLE with cutaneous manifestations is described by Slimani et.al. (2021). Autoantibodies are very commonly found in COVID-19 patients, including antibodies found in blood (Vlachoyiannopoulos et. al., 2020) and cerebrospinal fluid (CFS) (Franke et. al., 2021).
Though SARS-CoV-2 is not found in the CSF, it is theorized that the autoantibodies created in response to SARS-CoV-2 exposure may lead to at least some portion of the neurological complications documented in COVID-19 patients.
One important Letter to the Editor submitted to the journal Arthritis & Rheumatologyby Bertin et. al. (2020) noted the high prevalence and strong association (p=0.009) of autoantibodies against cardiolipin in COVID-19 patients with severe disease.
Zuo et. al. (2020) found anti-phospholipid autoantibodies in 52% of hospitalized COVID-19 patients and speculated that these antibodies contribute to the high incidence of coagulopathies in these patients.
Schiaffino et. al. (2020) reported that serum from a high percentage of hospitalized COVID-19 patients contained autoantibodies reactive to the plasma membrane of hepatocytes and gastric cells.
One patient with Guillain-Barre Syndrome was found to have antibody reactivity in cerebrospinal fluid (CFS), leading the authors to suggest that cross-reactivity with proteins in the CFS could lead to neurological complications seen in some COVID-19 patients.
In a more recent review, Gao et. al. (2021) noted high levels of autoantibodies in COVID-19 patients across multiple studies. They conclude, “[O]ne of the potential side effects of giving a mass vaccine could be an mergence [sic] of autoimmune diseases especially in individuals who are genetically prone for autoimmunity.”
A recent publication compiles a great deal of evidence that autoantibodies against a broad range of receptors and tissue can be found in individuals who have had previous SARS-CoV-2 infection. “All 31 former COVID-19 patients had between 2 and 7 different GPCR-fAABs [G-protein coupled receptor functional autoantibodies] that acted as receptor agonists.”(Wallukat et. al. 2021)
The diversity of GPCR-fAABs identified, encompassing both agonist and antagonist activity on target receptors, strongly correlated with a range of post-COVID-19 symptoms, including tachycardia, bradycardia, alopecia, attention deficit, PoTS, neuropathies, and others.
The same study, referencing the autoantibodies predicted by Lyons-Weiler (2020) mentioned above, notes with obvious grave concern: “The Sars-CoV-2 spike protein is a potential epitopic target for biomimicry-induced autoimmunological processes [25]. Therefore, we feel it will be extremely important to investigate whether GPCR-fAABs will also become detectable after immunisation by vaccination against the virus.”
We have reviewed the evidence here that the spike protein of SARS-CoV-2 has extensive sequence homology with multiple endogenous human proteins and could prime the immune system toward development of both auto-inflammatory and autoimmune disease.
This is particularly concerning given that the protein has been redesigned with two extra proline residues to potentially impede its clearance from the circulation through membrane fusion.
These diseases could present acutely and over relatively short timespans such as with MIS-C or could potentially not manifest for months or years following exposure to the spike protein, whether via natural infection or via vaccination.
Many who test positive for COVID-19 express no symptoms. The number of asymptomatic, PCR-positive cases varies widely between studies, from a low of 1.6% to a high of 56.5% (Gao et. al., 2020).
Those who are insensitive to COVID-19 probably have a very strong innate immune system. The healthy mucosal barrier’s neutrophils and macrophages rapidly clear the viruses, often without the need for any antibodies to be produced by the adaptive system. However, the vaccine intentionally completely bypasses the mucosalimmune system, both through its injection past the natural mucosal barriers and its artificial configuration as an RNA-containing nanoparticle.
As noted in Carsetti (2020), those with a strong innate immune response almost universally experience either asymptomatic infection or only mild COVID-19 disease presentation. Nevertheless, they might face chronic autoimmune disease, as described previously, as a consequence of excessive antibody production in response to the vaccine, which was not necessary in the first place.
The Spleen, Platelets and Thrombocytopenia
Dr. Gregory Michael, an obstetrician in Miami Beach, died of a cerebral hemorrhage 16 days after receiving the first dose of the Pfizer/BioNTech COVID-19 vaccine. Within three days of the vaccine, he developed idiopathic thrombocytopenic purpura (ITP), an autoimmune disorder in which the immune cells attack and destroy the platelets. His platelet count dropped precipitously, and this caused an inability to stop internal bleeding, leading to the stroke, as described in an article in the New York Times (Grady and Mazzei, 2021).
The New York Times followed up with a second article that discussed several other cases of ITP following SARS-CoV-2 vaccination (Grady, 2021), and several other incidences ofprecipitous drop of platelets and thrombocytopenia following SARS-CoV-2 vaccination have been reported in the Vaccine Adverse Event Reporting System (VAERS).
1. Biodistribution of mRNA Vaccines
Several studies on mRNA-based vaccines have confirmed independently that the spleen is a major center of activity for the immune response. A study on an mRNA-based influenza virus vaccine is extremely relevant for answering the question of the biodistribution of the mRNA in the vaccine.
This vaccine, like the SARS-CoV-2 vaccines, was designed as lipid nanoparticles with modified RNA coding for hemagglutinin (the equivalent surface fusion protein to the spike protein in corona viruses), and was administered through muscular injection.
The concentration of mRNA was tracked over time in various tissue samples, and the maximum concentration observed at each site was recorded. Not surprisingly, the concentration was highest in the muscle at the injection site (5,680 ng/mL).
This level decreased slowly over time, reaching half the original value at 18.8 hours following injection. The next highest level was observed in the proximal lymph node, peaking at 2,120 ng/mL and not dropping to half this value until 25.4 hours later. Among organs, the highest levels by far were found in the spleen (86.69 ng/mL) and liver (47.2 ng/mL). Elsewhere in the body the concentration was at 100-to 1,000-fold lower levels. In particular, distal lymph nodes only had a peak concentration of 8 ng/mL.
They concluded that the mRNA distributes from the injection site to the liver and spleen via the lymphatic system, ultimately reaching the general circulation. This likely happens through its transport inside macrophages and other immune cells that take it up at the muscular injection site.
Disturbingly, it also reaches into the brain, although at much lower levels (Bahl et al., 2017).
The European Medicines Agency assessment report for the Moderna vaccine also noted that mRNA could be detected in the brain following intramuscular administration at about 2% of the level found in the plasma (European Medicines Agency, 2021).
In another experiment conducted to track the biodistribution pathway of RNA vaccines, a rabies RNA vaccine was administered intramuscularly to rats in a single dose. The vaccine included a code for an immunogenic rabies protein as well as the code for RNA polymerase and was formulated as an oil-in-water nanoemulsion.
Thus, it is not entirely representative of the SARS-CoV-2 mRNA vaccines. Nevertheless, its intramuscular administration and its dependence on RNA uptake by immune cells likely means that it would migrate through the tissues in a similar pathway as the SARS-CoV-2 vaccine.
The authors observed an enlargement of the draining lymph nodes, and tissue studies revealed that the rabies RNA appeared initially at the injection site and in the draining lymph nodes within one day, and was also found in blood, lungs, spleen and liver (Stokes et al., 2020).
These results are consistent with the above study on influenza mRNA vaccines.Finally, a study comparing luciferase-expressing mRNA nanoparticles with luciferase-expressing mRNA dendritic cells as an alternative approach to vaccination revealed that the luciferase signal reached a broader range of lymphoid sites with the nanoparticle delivery mechanism.
More importantly, the luciferase signal was concentrated in the spleen for the nanoparticles compared to dominance in the lungs for the dendritic cells (Firdessa-Fite and Creuso, 2020).
2.Immune Thrombocytopenia
Immune thrombocytopenia (ITP) has emerged as an important complication of COVID-19 (Bhattacharjee and Banerjee, 2020). In many cases, it emerges after full recovery from the disease, i.e, after the virus has been cleared, suggesting it is an autoimmune phenomenon.
A likelypathway by which ITP could occur following vaccination is through the migration of immune cells carrying a cargo of mRNA nanoparticles via the lymph system into the spleen. These immune cells would produce spike protein according to the code in the nanoparticles, and the spike protein would induce B cell generation of IgG antibodies to it.
ITP appears initially as petechiae or purpura on the skin, and/or bleeding from mucosal surfaces. It has a high risk of fatality through haemorrhaging and stroke. ITP is characterized by both increased platelet destruction and reduced platelet production, and autoantibodies play a pivotal role (Sun and Shan, 2019).
Platelets are coated by anti-platelet antibodies and immune complexes, and this induces their clearance by phagocytes. Particularly under conditions of impaired autophagy, the resulting signaling cascade can also result in suppression of production of megakaryocytes in the bone marrow, which are the precursor cells for platelet production (Sun and Shan, 2019).
A case study of a patient diagnosed with COVID-19 is revealing because he developed sudden onset thrombocytopenia a couple of days after he had been released from the hospital based on a negative COVID-19 nucleic acid test. Following this development, it was verified that the patient had a reduced number of platelet-producing megakaryocytes, while autoimmune antibodies were negative, suggesting a problem with platelet production rather than platelet destruction (Chen et al., 2020).
Platelets appear to play an important role in viral clearance. Within one minute after platelets were incubated together with influenza viruses, the viruses had already attached to the platelets. Subsequent internalization, possibly by phagocytosis, peaked at 30 minutes (Jansen et al., 2020).
The SARS-CoV-2 spike protein bindssialic acid, which means it could attach to glycoproteins in the platelet membranes (Baker et al., 2020). There is a structural similarity between the S1 spike protein in SARS CoV and neuraminidase expressed by the influenza virus, which might mean that the spike protein possesses neuraminidase activity (Zhang et al., 2004).
Several viruses express neuraminidase, and it generally acts enzymatically to catabolize the glycans in glycoproteins through desialylation. Thus, it seems plausible that a dangerous cascade leading to ITP could ensue following mRNA vaccination, even with no live virus present, particularly in the context of impaired autophagy.
Immune cells in the arm muscle take up the RNA particles and circulate within the lymph system, accumulating in the spleen. There, the immune cells produce abundant spike protein, which binds to the platelet glycoproteins and desialylates them. Platelet interaction with neutrophils causes NETosis and the launch of an inflammatory cascade.
The exposed glycoproteins become targets for autoimmune antibodies that then attack and remove the platelets, leading to a rapid drop in platelet counts, and a life-threatening event.
Activation of Latent Herpes Zoster
An observational study conducted at Tel Aviv Medical Center and the Carmel Medical Center in Haifa, Israel, found a significantly increased rate of herpes zoster following the Pfizer vaccination (Furer 2021). This observational study monitored patients with pre-existing autoimmune inflammatory rheumatic diseases (AIIRD). Among the 491 patients with AIIRD over the study period, 6 (1.2%) were diagnosed with herpes zoster as a first-ever diagnosis between 2 days and 2 weeks after either the first or second vaccination.
In the control group of 99 patients there were no herpes zoster cases identified. The CDC’s VAERS database, queried on April 19, 2021, contains 278 reports of herpes zoster following either the Moderna or Pfizer vaccinations.
Given the documented underreporting to VAERS (Lazarus et al. 2010), and given the associational nature of VAERS reports, it is not possible to prove any causal link between the vaccinations and the zoster reports.
However, we believe the occurrence of zoster is another important ‘signal’ in VAERS.This increased risk to shingles, if valid, may have important broader implications. Multiple studies have shown that patients with either primary or acquired immune deficiency are more susceptible to severe herpes zoster infection (Ansari et al., 2020).
This suggests that the mRNA vaccines may be suppressing the innate immune response. There is cross-talk between TNF-α and type I interferon in autoimmune disease, wherein each suppresses the other (Palucka et al., 2005). Type I interferon inhibits varicella-zoster virus replication (Ku et al., 2016). TNF-α is sharply upregulated in an inflammatory response, which is induced by the lipid nanoparticles in the vaccine. Its upregulation is also associated with the chronic inflammatory state of rheumatoid arthritis (Matsuno et al., 2002). Exuberant TNF-α expression following vaccination may be interfering with the dendritic cell INF-α response that keeps latent herpes zoster in check.
Spike Protein Toxicity
The picture is now emerging that SARS-CoV-2 has serious effects on the vasculature in multiple organs, including the brain vasculature. As mentioned earlier, the spike protein facilitates entry of the virus into a host cell by binding to ACE2 in the plasma membrane. ACE2 is a type I integral membrane protein that cleaves angiotensin II into angiotensin(1-7), thus clearing angiotensin II and lowering blood pressure. In a series of papers, Yuichiro Suzuki in collaboration with other authors presented a strong argument that the spike protein by itself can cause a signaling response in the vasculature with potentially widespread consequences (Suzuki, 2020; Suzuki et al., 2020; Suzuki et al., 2021; Suzuki and Gychka, 2021).
These authors observed that, in severe cases of COVID-19, SARS-CoV-2 causes significant morphological changes to the pulmonary vasculature. Post-mortem analysis of the lungs of patients who died from COVID-19 revealed histological features showing vascular wall thickening, mainly due to hypertrophy of the tunica media.
Enlarged smooth muscle cells had become rounded, with swollen nuclei and cytoplasmic vacuoles (Suzuki et al., 2020). Furthermore, they showed that exposure of cultured human pulmonary artery smooth muscle cells to the SARS-CoV-2 spike protein S1 subunit was sufficient to promote cell signaling without the rest of the virus components.
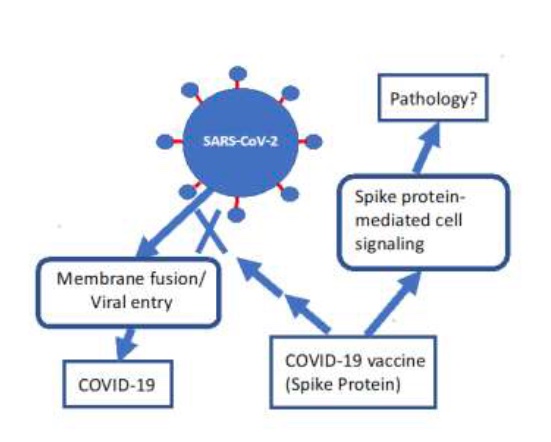
Figure 2: A simple model for a process by which the spike protein produced through the mRNA vaccines could induce a pathological response distinct from the desirable induction of antibodies to suppress viral entry. Redrawn with permission from Suzuki and Gychka, 2021.
Follow-on papers (Suzuki et al., 2021, Suzuki and Gychka, 2021) showed that the spike protein S1 subunit suppresses ACE2, causing a condition resembling pulmonary arterial hypertension (PAH), a severe lung disease with very high mortality. Their model is depicted here in Figure 2.
Ominously, Suzuki and Gychka (2021) wrote: “Thus, these in vivo studies demonstrated that the spike protein of SARS-CoV-1 (without the rest of the virus) reduces the ACE2 expression, increases the level of angiotensin II, and exacerbates the lung injury.”
The “in vivo studies” they referred to here (Kuba et al., 2005) had shown that SARS coronavirus-induced lung injury was primarily due to inhibition of ACE2 by the SARS-CoV spike protein, causing a large increase in angiotensin-II.
Suzuki et al. (2021) went on to demonstrate experimentally that the S1 component of the SARS-CoV-2 virus, at a low concentration of 130 pM, activated the MEK/ERK/MAPK signaling pathway to promote cell growth. They speculated that these effects would not be restricted to the lung vasculature.
The signaling cascade triggered in the heart vasculature would cause coronary artery disease, and activation in the brain could lead to stroke. Systemic hypertension would also be predicted. They hypothesized that this ability of the spike protein to promote pulmonary arterial hypertension could predispose patients who recover from SARS-CoV-2 to later develop right ventricular heart failure.
Furthermore, they suggested that a similar effect could happen inresponse to the mRNA vaccines, and they warned of potential long-term consequences to both children and adults who received COVID-19 vaccines based on the spike protein (Suzuki and Gychka, 2021).
An interesting study by Lei et. al. (2021) found that pseudovirus —spheres decorated with the SARS-CoV-2 S1 protein but lacking any viral DNA in theircore —caused inflammation and damage in both the arteries and lungs of mice exposed intratracheally.
They then exposed healthy human endothelial cells to the samepseudovirus particles. Binding of these particles to endothelial ACE2 receptors led to mitochondrial damage and fragmentation in those endothelial cells, leading to the characteristic pathological changes in the associated tissue.
This study makes it clear that spike protein alone, unassociated with the rest of the viral genome, is sufficient to cause the endothelial damage associated with COVID-19.
The implications for vaccines intended to cause cells to manufacture the spike protein are clear and are an obvious cause for concern.
Neurological symptoms associated with COVID-19, such as headache, nausea and dizziness, encephalitis and fatal brain blood clots are all indicators of damaging viral effects on the brain.
Buzhdygan et al. (2020) proposed that primary human brain microvascular endothelial cells could cause these symptoms. ACE2 is ubiquitously expressed in the endothelial cells in the brain capillaries. ACE2 expression is upregulated in the brain vasculature in association with dementia and hypertension, both of which are risk factors for bad outcomes from COVID-19.
In an in vitro study of the blood-brain barrier, the S1 component of the spike protein promoted loss of barrier integrity, suggesting that the spike protein acting alone triggers a pro-inflammatory response in brain endothelial cells, which could explain the neurological consequences of the disease (Buzhdygan et al., 2020).
The implications of this observation are disturbing because the mRNA vaccines induce synthesis of the spike protein, which could theoretically act in a similar way to harm the brain.
The spike protein generated endogenously by the vaccine could also negatively impact the male testes, as the ACE2 receptor is highly expressed in Leydig cells in the testes (Verma et al., 2020). Several studies have now shown that the coronavirus spike protein is able to gain access to cells in the testes via the ACE2 receptor, and disrupt male reproduction (Navarra et al., 2020; Wang and Xu, 2020). A paper involving postmortem examination of testicles of six male COVID-19 patients found microscopic evidence of spike protein in interstitial cells in the testes of patients with damaged testicles (Achua et al., 2021).
A Possible Link to Prion Diseases and Neurodegeneration
Prion diseases are a collection of neurodegenerative diseases that are induced through the misfolding of important bodily proteins, which form toxic oligomers that eventually precipitate out as fibrils causing widespread damage to neurons.
Stanley Prusiner first coined the name `prion’ to describe these misfolded proteins (Prusiner, 1982). The best-known prion disease is MADCOW disease (bovine spongiform encephalopathy), which became an epidemic in European cattle beginning in the 1980s.
The CDC web site on prion diseases states that “prion diseases are usually rapidly progressive and always fatal.”(Centers for Disease Control and Prevention, 2018). It is now believed that many neurodegenerative diseases, including Alzheimer’s, Parkinson’s disease, and amyotrophic lateral sclerosis (ALS) may be prion diseases, and researchers have identified specific proteinaceous infectious particles linked to these diseases (Weickenmeier et al., 2019).
Furthermore, researchers have identified a signature motif linked to susceptibility to misfolding into toxic oligomers, called the glycine zipper motif. It is characterized by a pattern of two glycine residues spaced by three intervening amino acids, represented as GxxxG.
The bovine prion linked to MADCOW has a spectacular sequence of ten GxxxGs in a row (see uniprot.org/uniprot/P10279).
More generally, the GxxxG motif is a common feature of transmembrane proteins, and the glycines play an essential role in cross-linking α-helices in the protein (Mueller et al., 2014).
Prion proteins become toxic when the α-helices misfold as β-sheets, and the protein is then impaired in its ability to enter the membrane (Prusiner, 1982).
Glycines within the glycine zipper transmembrane motifs in the amyloid-β precursor protein (APP) play a central role in the misfolding of amyloid-β linked to Alzheimer’s disease (Decock et al., 2016).
APP contains a total of four GxxxG motifs. When considering that the SARS-CoV-2 spike protein is a transmembrane protein, and that it contains fiveGxxxG motifs in its sequence (see uniprot.org/uniprot/P0DTC2), it becomes extremely plausible that it could behave as a prion.
One of the GxxxG sequences is present within its membrane fusion domain. Recall that the mRNA vaccines are designed with an altered sequencethat replaces two adjacent amino acids in the fusion domain with a pair of prolines.
This is done intentionally in order to force the protein to remain in its open state and make it harder for it to fuse with the membrane. This seems to us like a dangerous step towards misfolding potentially leading to prion disease.
A paper published by J. Bart Classen (2021) proposed that the spike protein in the mRNA vaccines could cause prion-like diseases, in part through its ability to bind to many known proteins andinduce their misfolding into potential prions. Idrees and Kumar (2021) have proposed that the spike protein’s S1 component is prone to act as a functional amyloid and form toxic aggregates.
These authors wrote that S1 has the ability “to form amyloid and toxic aggregates that can act as seeds to aggregate many of the misfolded brain proteins and can ultimately lead to neurodegeneration.”According to Tetz and Tetz (2020), the form of the spike protein in SARS-CoV-2 has prion regions that are not present inthe spike proteins for other coronaviruses. While this was reported in a non-peer-reviewed article, the authors had published a previous paper in 2018 identifying prion-like regions in multiple eukaryotic viruses, so they have considerable expertise in this area (Tetz and Tetz, 2018).
A final point here relates to information about the Pfizer vaccine in particular. The European Medicines Agency (EMA) Public Assessment Report is a document submitted to gain approval to market the vaccine in Europe. It describes in detail a review of the manufacturing process as well as a wide range of associated testing data.
One concerning revelation is the presence of “fragmented species” of RNA in the injection solution. These are RNA fragments resulting from early termination of the process of transcription from the DNA template. These fragments, if translated by the cell following injection, would generate incomplete spike proteins, again resulting in altered and unpredictable three-dimensional structure and a physiological impact that is at best neutral and at worst detrimental to cellular functioning.
There were considerably more of these fragmented forms of RNA found in the commercially manufactured products than in the products used in clinical trials. The latter were produced via a much more tightly controlled manufacturing process.
Pfizer claims the RNA fragments “likely… will not result in expressed proteins” due to their assumed rapid degradation within the cell. No data was presented to rule out protein expression, though, leaving the reviewers to comment, “These [fragmented RNA] forms are poorly characterised, and the limited data provided for protein expression do not fully address the uncertainties relating to the risk of translating proteins/peptides other than the intended spike protein” (EMA 2020).
To our knowledge no data has been forthcoming since that time. While we are not asserting that non-spike proteins generated from fragmented RNA would be misfolded or otherwise pathological, we believe they would at least contribute to the cellular stress that promotes prion-associated conformational changes in the spike protein that is present.
1.Lessons from Parkinson’s Disease
Parkinson’s disease is a neurodegenerative disease associated with Lewy body deposits in the brain, and the main protein found in these Lewy bodies is α-synuclein. That protein, α-Synuclein, is certainly prion-like insofar as under certain conditions it aggregates into toxic soluble oligomers and fibrils (Lema Tomé et al., 2013).
Research has shown that misfolded α-synuclein can form first in the gut and then travel from there to the brain along the vagus nerve, probably in the form of exosomes released from dying cells where the misfolded protein originated (Kakarla et al., 2020; Steiner et al., 2011).
The cellular conditions that promote misfolding include both an acidic pH and high expression of inflammatory cytokines.
It is clear that the vagus nerve is critical for transmission of misfolded proteins to the brain, because severance of the vagus nerve protects from Parkinson’s. Vagus nerve atrophy in association with Parkinson’s disease provides further evidence of the involvement of the vagus nerve in transport of misfolded α-synuclein oligomers from the gut to the brain (Walter et al., 2018).
Another pathway is through the olfactory nerve, and a loss of a sense of smell is an early sign of Parkinson’s disease. Ominously, diminution or loss of the sense of smell is also a common symptom of SARS-CoV-2 infection. There are many parallels between α-synuclein and the spike protein, suggesting the possibility of prion-like disease following vaccination.
We have already shown that the mRNA in the vaccine ends up in high concentrations in the liver and spleen, two organs that are well connected to the vagus nerve.
The cationic lipids in the vaccine create an acidic pH conducive to misfolding, and they also induce a strong inflammatory response, another predisposing condition.
Germinal centers are structures within the spleen and other secondary lymphoid organs where follicular dendritic cells present antigens to Bcells, which in turn perfect their antibody response. Researchers have shown that mRNA vaccines, in contrast with recombinant protein vaccines, elicit a robust development of neutralizing antibodies at these germinal centers in the spleen (Lederer et al., 2020).
However, this also means that mRNA vaccines induce an ideal situation for prion formation from the spike protein, and its transport via exosomes along the vagus nerve to the brain.
Studies have shown that prion spread from one animal to another first appears in the lymphoid tissues, particularly the spleen. Differentiated follicular dendritic cells are central to the process, as they accumulate misfolded prion proteins (Al-Dybiat et al., 2019).
An inflammatory response upregulates synthesis of α-synuclein in these dendritic cells, increasing the risk of prion formation. Prions that accumulate in the cytoplasm are packaged up into lipid bodies that are released as exosomes (Liu et al., 2017).
These exosomes eventually travel to the brain, causing disease. 2.Vaccine SheddingThere has been considerable chatter on the Internet about the possibility of vaccinated people causing disease in unvaccinated people in close proximity. While this may seem hard to believe, there is a plausible process by which it could occur through the release of exosomes from dendritic cells in the spleen containing misfolded spike proteins, in complex with other prion reconformed proteins. These exosomescan travel to distant places. It is not impossible to imagine that they are being released from the lungs and inhaled by a nearby person. Extracellular vesicles, including exosomes, have been detected in sputum, mucus, epithelial lining fluid, and bronchoalveolar lavage fluid in association with respiratory diseases (Lucchetti et al., 2021).
A Phase 1/2/3 study undertaken by BioNTech on the Pfizer mRNA vaccine implied in their study protocol that they anticipated the possibility of secondary exposure to the vaccine (BioNTech, 2020).
The protocol included the requirement that “exposure during pregnancy” should be reported by the study participants. They then gave examples of “environmental exposure during pregnancy” which included exposure “to the study intervention by inhalation or skin contact.” They even suggested two levels of indirect exposure: “A male family member or healthcare provider who has been exposed to the study intervention by inhalation or skin contact then exposes his female partner prior toor around the time of conception.”
Emergence of Novel Variants of SARS-CoV-2
An interesting hypothesis has been proposed in a paper published in Nature, which described a case of serious COVID-19 disease in a cancer patient who was taking immune-suppressing cancer chemotherapy drugs (Kemp et al., 2021).
The patient survived for 101 days after admission to the hospital, finally succumbing in the battle against the virus. The patient constantly shed viruses over the entire 101 days, and therefore he was moved to a negative-pressure high air-change infectious disease isolation room, to prevent contagious spread.
During the course of the hospital stay, the patient was treated with Remdesivir and subsequently with two rounds of antibody-containing plasma taken from individuals who had recovered from COVID-19 (convalescent plasma).
It was only after the plasma treatments that the virus began to rapidly mutate, and a dominant new strain eventually emerged, verified from samples taken from the nose and throat of the patient.
An immune-compromised patient offers little support from cytotoxic T cells to clear the virus. An in vitro experiment demonstrated that this mutant strain had reduced sensitivity to multiple units of convalescent plasma taken from several recovered patients.
The authors proposed that the administered antibodies had actually accelerated the mutation rate in the virus, because the patient was unable to fully clear the virus due to their weak immune response. This allowed a “survival of the fittest” program to set in, ultimately populating the patient’s body with a novel antibody-resistant strain. Prolonged viral replication in this patient led to “viral immune escape,” and similar resistant strains could potentially spread very quickly within an exposed population (Kemp et al., 2021).
Indeed, a similar process might plausibly be at work to produce the highly contagious new strains that are now appearing in the United Kingdom, South Africa and Brazil. There are at least two concerns that we have regarding this experiment, in relation to the mRNA vaccines.
The first is that, via continued infection of immune-compromised patients, we can expect continued emergence of more novel strains that are resistant to the antibodies induced by the vaccine, such that the vaccine may quickly become obsolete, and there may well be demands for the population to undergo another mass vaccination campaign.
Already a published study by researchers from Pfizer has shown that vaccine effectiveness is reduced for many of these variant strains. The vaccine was only 2/3 as effective against the South African strain as against the original strain (Liu et al., 2021).
The second more ominous consideration is to ponder what will happen with an immune-compromised patient following vaccination. It is conceivable that they will respond to the vaccine by producing antibodies, but those antibodies will be unable to contain the disease following exposure to COVID-19 due to impaired function of cytotoxic T cells.
This scenario is not much different from the administration of convalescent plasma to immune-compromised patients, and so it might engender the evolution of antibody-resistant strains in the same way, only on a much grander scale. This possibility will surely be used to argue for repeated rounds of vaccines every few months, with increasing numbers of viral variants coded into the vaccines. This is an arms race that we will probably lose.
Potential for Permanent Incorporation of Spike Protein Gene into human DNA
It has been claimed that mRNA-based vaccines are safer than DNA-vectored vaccines that work by incorporating the genetic code for the target antigenic protein into a DNA virus, because the RNA cannot become inadvertently incorporated into the human genome.
However, it is not at all clear that this is true. The classic model of DNA → RNA → protein is now known to be false. It is now indisputable that there is a large class of viruses called retroviruses that carry genes that reverse transcribe RNA back into complementary DNA (cDNA).
In 1975, Howard Temin, Renato Dulbecco, and David Baltimore shared the Nobel Prize in Physiology or Medicine in 1975 for their discovery of reverse transcriptase and its synthesis by retroviruses (such as human immunodeficiency virus (HIV)) to derive DNA from RNA (Temin and Mizutani, 1970, Baltimore, 1970).
Much later, it was discovered that reverse transcriptase is not unique to retroviruses. More than a third of the human genome is devoted to mysterious mobile DNA elements called SINEs and LINEs (short and long interspersed nuclear elements, respectively).
LINEs provide reverse transcriptase capabilities to convert RNA into DNA, and SINEs provide support for integrating the DNA into the genome. Thus, these elements provide the tools needed to convert RNA into DNA and incorporate it into the genome so as to maintain the new gene through future generations (Weiner, 2002).
SINEs and LINEs are members of a larger class of genetic elements called retrotransposons. Retrotransposons can copy and paste their DNA to a new site in the genome via an RNA intermediate, while possibly introducing genetic alterations in the process (Pray, 2008).
Retrotransposons, also known as “jumping genes,” were first identified by the geneticist Barbara McClintock of ColdSpring Harbor Laboratory in New York, over 50 years ago (McClintock, 1965).
Much later, in 1983, she was recognized with a Nobel prize for this work. Remarkably, retrotransposons seem to be able to expand their domain from generation to generation. LINEs and SINEs collaborate to invade new genomic sites through translation of their DNA to RNA and back to a fresh copy of DNA, which is then inserted at an AT-rich region of the genome.
These LINEs and SINEs had long been considered to be “junk” DNA, an absurd idea that has now been dispelled, as awareness of their critical functions has grown.
In particular, it has now become clear that they can also import RNA from an exogenous source into a mammalian host’s DNA. Retroviral-like repeat elements found in the mouse genome called intracisternal A particles (IAPs) have been shown to be capable of incorporating viral RNA into the mouse genome.
Recombination between an exogenous nonretroviral RNA virus and an IAP retrotansposon resulted in reverse transcription of the viral RNA and integration into the host’s genome (Geuking et al., 2009).
Furthermore, as we shall see later, the mRNA in the new SARS-CoV-2 vaccines could also get passed on from generation to generation, with the help of LINEs expressed in sperm, vianon-integrated cDNA encapsulated in plasmids. The implications of this predictable phenomenon are unclear, but potentially far-reaching.
1. Exogenous and Endogenous Retroviruses
There is also a concern that the RNA in the mRNA vaccines could be transferred into the human genome with assistance from retroviruses.
Retroviruses are a class of viruses that maintain their genomic information in the form of RNA, but that possess the enzymes needed to reverse transcribe their RNA into DNA and insert it into a host genome. They then rely on existing natural tools from the host to produce copies of the virus through translation of DNA back into RNA and to produce the proteins that the viral RNA codes for and assemble them into a fresh viral particle (Lesbats et al., 2016).
Human endogenous retroviruses (HERVs) are benign sections in the DNA of humans that closely resemble retroviruses, and that are believed to have become permanent sequences in the human genome through a process of integration from what was originally an exogenous retrovirus.
Endogenous retroviruses are abundant in all jawed vertebrates and are estimated to occupy 5-8% of the human genome. The protein syncytin, which has become essential for placental fusion with the uterine wall and for the fusion step between the sperm and the egg at fertilization, is a good example of an endogenous retroviral protein.
Syncytin is the envelope gene of a recently identified human endogenous defective retrovirus, HERV-W (Mi et al., 2000). During gestation, the fetus expresses high levels of another endogenous retrovirus, HERV-R, and it appears to protect the fetus from immune attack from the mother (Luganini and Gribaudo, 2020).
Endogenous retroviral elements closely resemble retrotransposons. Their reverse transcriptase, when expressed, has the theoretical capability to convert spike protein RNA from the mRNA vaccines into DNA.
2. Permanent DNA integration of Exogenous Retrovirus Genes
Humans are colonized by a large collection of exogenous retroviruses that in many cases cause no harm to the host, and may even be symbiotic (Luganini and Gribaudo, 2020).
Exogenous viruses can be converted to endogenous viruses (permanently incorporated into host DNA) in the laboratory, as demonstrated by Rudolf Jaenisch (Jaenisch, 1976), who infected preimplantation mouse embryos with the Moloney murine leukemia virus (M-MuLV).
The mice generated from these infected embryos developed leukemia, and the viral DNA was integrated into their germ line and transmitted to their offspring. Besides the incorporation of viral DNA into the host genome, it was also shown as early as 1980 that DNA plasmids could be microinjected into the nuclei of mouse embryos to produce transgenic mice that breed true (Gordon et al., 1980).
The plasmid DNA was incorporated into the nuclear genome of the mice through existing natural processes, thus preserving the newly acquired genetic information in the offspring’s genome. This discovery has been the basis for many genetic engineering experiments on transgenic mice engineered to express newly acquired human genes since then (Bouabe and Okkenhaug, 2013).
3. LINE-1 is Widely Expressed
LINEs alone make up over 20% of the human genome. The most common LINE is LINE-1, which encodes a reverse transcriptase that regulates fundamental biological processes. LINE-1 is expressed in many cell types, but at especially high levels in sperm.
Sperm cells can be used as vectors of both exogenous DNA and exogenous RNA molecules through sperm-mediated gene transfer assays. Sperm can reverse transcribe exogenous RNA directly into cDNA and can deliver plasmids packaging up this cDNA to the fertilized egg.
These plasmids are able to propagate themselves within the developing embryo and to populate many tissues in the fetus. In fact, they survive into adulthood as extrachromosomal structures and are capable of being passed on to progeny.
These plasmids are transcriptionally competent, meaning that they can be used to synthesize proteins encoded by the DNA they contain (Pittoggi et al., 2006).
In addition to sperm, embryos also express reverse transcriptase prior to implantation, and its inhibition causes developmental arrest. LINE-1 is also expressed by cancer cells, and RNA interference-mediated silencing of human LINE-1 induces differentiation in many cancer cell lines.
Reverse-transcriptase machinery is implicated in the genesis of new genetic information, both in cancer cells and in germ cells. Many tumor tissues have been found to express high levels of LINE-1, and to contain many extrachromosomal plasmids in their nucleus.
Malignant gliomas are the primary tumors of the central nervous system. It has beenshown experimentally that these tumors release exosomes containing DNA, RNA and proteins, that end up in the general circulation (Vaidya and Sugaya, 2020).
LINE-1 is also highly expressed in immune cells in several autoimmune diseases such as systemic lupus erythematosus, Sjögrens and psoriasis (Zhang et al., 2020).
4. Integrating Spike Protein Gene into Human Genome
Remarkably, it has been demonstrated that neurons from the brain of Alzheimer’s patients harbor multiple variants of the gene for amyloid precursor protein APP, incorporated into the genome, which are created through a process called somatic gene recombination (SGR) (Kaeser et al., 2020).
SGR requires gene transcription, DNA strand-breaks, and reverse transcriptase activity, all of which may be promoted by well-known Alzheimer’s disease risk factors.
The DNA coding for APP is reverse transcribed into RNA and then transcribed back into DNA and incorporated into the genome at a strand break site. Since RNA is more susceptible to mutations, the DNA in these mosaic copies contains many mutant variants of the gene, so the cell becomes a mosaic, capable of producing multiple variants of APP.
Neurons from Alzheimer’s patients contained as many as 500 million base pairs of excess DNA in their chromosomes (Bushman et al., 2015).
Researchers from MIT and Harvard published a disturbing paper in 2021, where they provided strong evidence that the SARS-CoV-2 RNA can be reverse transcribed into DNA and integrated into human DNA (Zhang et al., 2021).
They were led to investigate this idea after having observed that many patients continue to test positive for COVID-19 after the virus has already been cleared from their body. The authors found chimeric transcripts that contained viral DNA sequences fused to cellularDNA sequences in patients who had recovered from COVID-19.
Since COVID-19 often induces a cytokine storm in severe cases, they confirmed the possibility of enhanced reverse transcriptase activity through an in vitro study using cytokine-containing conditioned media in cell cultures. They found a 2-3-fold upregulation of endogenous LINE-1 expression in response to cytokines. The exogenous RNA from the virus incorporated into human DNA could produce fragments of viral proteins indefinitely after the infection has been cleared, and this yields a false-positive on a PCR test.
5. Bovine Viral Diarrhea: A Disturbing Model
Bovine Viral Diarrhea (BVD) is an infectious viral disease that affects cattle throughout the world. It is a member of the class of pestiviruses, which are small, spherical, single-stranded, enveloped RNA viruses.
The disease is associated with gastrointestinal, respiratory and reproductive diseases. A unique characteristic of BVD is that the virus can cross the placenta of an infected pregnant dam. This can result in the birth of a calf which carries intra-cellular viral particles which it mistakes as `self.’ Its immune system refuses to recognize the virus as a foreign invasion, and, as a result, the calf sheds the virus in large quantities throughout its life, potentially infecting the entire herd.
It has become a widespread practice to identify such carrier calves and cull them from the herd in an attempt to curtail infection (Khodakaram-Tafti & Farjanikish, 2017).
It seems plausible that a dangerous situation may arise in the future where a woman receives an mRNA vaccine for SARS-CoV-2 and then conceives a child shortly thereafter. The sperm would be free to take up RNA-embedded liposomes from the vaccine and convert them to DNA using LINE-1.
They would then produce plasmids containing the code for the spike protein which would be taken up by the fertilized egg through the process described above. The infant that is born is then potentially unable to mount antibodies to the spike protein because their immune system considers it to be `self.’
Should that infant get infected with SARS-CoV-2 at any time in its lifespan, its immune system would not mount a defense against the virus, and the virus would presumably be free to multiply in the infant’s body without restraint. The infant would logically become a super-spreader in such a situation.
Admittedly, this is speculation at this time, but there is evidence from what we know about retrotransposons, sperm, fertilization, the immune system and viruses, that such a scenario cannot be ruled out.
It has already been demonstrated in mouse experiments that the genetic elements in DNA vector vaccines, which are essentially plasmids, can integrate into the host genome (Wang et al., 2004). In fact, such a process has been suggested as a basis for Lamarckian evolution defined as the inheritance of acquired traits (Steele, 1980).
The realization that what was formerly called “junk DNA” is not junk, is just one of the results coming out of the new philosophical paradigm in human language, biology and genetics that is based on fractal genomics (Pellionisz, 2012) —a paradigm that Pellionisz has linked to the involvement of “true narrative representations” (TNRs; Oller, 2010), realized as “iterations of a fractal template” in the highly repetitive processes of normal development of the many branching structures of the human body.
These processes are numerous in the lungs, kidneys, veins and arteries, and most importantly in the brain. The mRNA vaccines are an experimental gene therapy with the potential to incorporate the code for the SARS-CoV-2 spike protein into human DNA.
This DNA code could instruct the synthesis of large numbers of copies of proteinaceous infectious particles, and this has the potential to insert multiple false signals into the unfolding narrative, resulting in unpredictable outcomes.
Conclusion
Experimental mRNA vaccines have been heralded as having the potential for great benefits, but they also harbor the possibility of potentially tragic and even catastrophic unforeseen consequences.
The mRNA vaccines against SARS-CoV-2 have been implemented with great fanfare, but there are many aspects of their widespread utilization that merit concern. We have reviewed some, but not all, of those concerns here, and we want to emphasize that these concerns are potentially serious and might not be evident for years or even transgenerationally.
In order to adequately rule out the adverse potentialities described in this paper, we recommend, at a minimum, that the following research and surveillance practices be adopted:
•A national effort to collect detailed data on adverse events associated with the mRNA vaccines with abundant funding allocation, tracked well beyond the first couple of weeks after vaccination.
•Repeated autoantibody testing of the vaccine-recipient population. The autoantibodies tested could be standardized and should be based upon previously documented antibodies and autoantibodies potentially elicited by the spike protein. These include autoantibodies against phospholipids, collagen, actin, thyroperoxidase (TPO), myelin basic protein, tissue transglutaminase, and perhaps others.
•Immunological profiling related to cytokine balance and related biological effects. Tests should include, at a minimum, IL-6, INF-α, D-dimer, fibrinogen, and C-reactive protein.
•Studies comparing populations who were vaccinated with the mRNA vaccines and those who were not to confirm the expected decreased infection rate and milder symptoms of the vaccinated group, while at the same time comparing the rates of various autoimmune diseases and prion diseases in the same two populations.
•Studies to assess whether it is possible for an unvaccinated person to acquire vaccine-specific forms of the spike proteins from a vaccinated person in close proximity.
•In vitro studies to assess whether the mRNA nanoparticles can be taken up by sperm and converted into cDNA plasmids.
•Animal studies to determine whether vaccination shortly before conception can result in offspring carrying spike-protein-encoding plasmids in their tissues, possibly integrated into their genome.
•In vitro studies aimed to better understand the toxicity of the spike protein to the brain, heart, testes, etc. Public policy around mass vaccination has generally proceeded on the assumption that the risk/benefit ratio for the novel mRNA vaccines is a “slam dunk.” With the massive vaccination campaign well underway in response to the declared international emergency of COVID-19, we have rushed into vaccine experiments on a worldwide scale. At the very least, we should take advantage of the data that are available from these experiments to learn more about this new and previously untested technology. And, in the future, we urge governments to proceed with more caution in the face of new biotechnologies.
Finally, as an obvious but tragically ignored suggestion, the government should also be encouraging the population to take safe and affordable steps to boost their immune systems naturally, such as getting out in the sunlight to raise vitamin D levels (Ali, 2020), and eating mainly organic whole foods rather than chemical-laden processed foods (Rico-Campà et al., 2019).
Also, eating foods that are good sources of vitamin A, vitamin C and vitamin K2 should be encouraged, as deficiencies in these vitamins are linked to bad outcomes from COVID-19 (Goddek, 2020; Sarohan, 2020).
Acknowledgements
This research was funded in part by Quanta Computers, Inc., Taiwan, under the auspices of the Qmulus project.
Competing interests
The authors have no competing interests or conflicts to declare.
References
Achua, J. K., Chu, K. Y., Ibrahim, E., Khodamoradi, K., Delma, K. S., Ramsamy, R. … Arora, H. (2021). Histopathology and Ultrastructural Findings of Fatal COVID-19 Infections on Testis. The World Journal of Men’s Health 39(1): 65-74. https://doi.org/10.5534/wjmh.200170.
Al-Dybiat, I., Moudjou, M., Martin, D., Reine, F., Herzog, L., Truchet, S., … Sibille, P. (2019) Prion Strain-dependent Tropism is Maintained between Spleen and Granuloma and Relies on Lymphofollicular Structures. Scientific Reports 9: 14656. https://doi.org/10.1038/s41598-019-51084-1.
Ali, N. (2020). Role of Vitamin D in Preventing of COVID-19 Infection, Progression and Severity. Journal of Infection and Public Health 13(10): 1373-1380. https://doi.org/10.1016/j.jiph.2020.06.021.
Ansari, B. Rosen, L. B., Lisco, A., Gilden, D., Holland, S. M., Zerbe, C. S., … Cohen, J. I. (2020). Primary and Acquired Immunodeficiencies Associated with Severe Varicella-Zoster Virus Infections. Clinical Infectious Diseases August 28 [Epub ahead of print]. https://doi.org/10.1093/cid/ciaa1274.
Arvin, A. M., Fink, K. Schmid, M. A., Cathcart, A., Spreafico, R., Havenar-Daughton, C. … Virgin, H. W. (2020). A Perspective on Potential Antibody-Dependent Enhancement of SARS-CoV-2. Nature 584(7821): 353-363. https://doi.org/10.1038/s41586-020-2538-8.
Aslam, R., Kapur, R., egel, G. B., Guo, L., Zufferey, A., Ni, H. & Semple, J. W. (2016). The Spleen Dictates Platelet Destruction, Anti-platelet Antibody Production, and Lymphocyte Distribution Patterns in a Murine Model of Immune Thrombocytopenia. Experimental Hematology 44(10): 924-930. https://doi.org/10.1016/j.exphem.2016.07.004.
Baden, L. R., El Sahly, H. M., Essink, B.,Kotloff, K., Frey, S., Novak, R. … Zaks, T. (2021). Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. The New England Journal of Medicine 384: 403-416. https://doi.org/10.1056/NEJMoa2035389.
Bahl, K., Senn, J. J., Yuzhakov, O., Bulychev, A., Brito, L. A., Hassett, K. J. … Ciaramella, G. (2017). Preclinical and Clinical Demonstration of Immunogenicity by mRNA Vaccines against H10N8 and H7N9 Influenza Viruses. Molecular Therapy 25(6): 1316-1327. http://dx.doi.org/10.1016/j.ymthe.2017.03.035.
Baker, A. N., Richards,S.-J., Guy, C. S., Congdon, T. R., Hasan, M., Zwetsloot, A. J., … Gibson, M. I. (2020). The SARS-COV-2 Spike Protein Binds Sialic Acids and Enables Rapid Detection in a Lateral Flow Point of Care Diagnostic Device. ACS Central Science 6(11): 2046-2052. https://doi.org/10.1021/acscentsci.0c00855.
Baltimore, D. (1970). Viral RNA-dependent DNA Polymerase: RNA-dependent DNA Polymerase in visions of RNA Tumor Viruses. Nature 226(5252): 1209-1211. https://doi.org/10.1038/2261209a0.
Bardina, S. V., Bunduc, P., Tripathi, S., Duehr, J., Frere, J. J., Brown, J. A. … Lim, J. K. (2017). Enhancementof Zika Virus Pathogenesis by Preexisting Antiflavivirus Immunity. Science 356(6334): 175-180. https://doi.org/10.1126/science.aal4365.
Beltramello, M., Williams, K. L., Simmons, C. P., Macagno, A., Simonelli, L., Ha Quyen, N. T. … Sallusto, F. (2010). The Human Immune Response to Dengue Virus is Dominated by Highly Cross-Reactive Antibodies Endowed with Neutralizing and Enhancing Activity. Cell Host Microbe 8(3): 271-83. https://doi.org/10.1016/j.chom.2010.08.007.
Bertin, D., Brodovitch, A., Beziane, A., Hug, S., Bouamri, A., Mege, J. L. … Bardin, N. (2020). Anticardiolipin IgG Autoantibody Level Is an Independent Risk Factor for COVID‐19 Severity.Arthritis & Rheumatology,72(11), 1953-1955. https://doi.org/10.1002/art.41409.
Bhattacharjee, S. & Banerjee, M. (2020). Immune Thrombocytopenia Secondary to COVID-19: a Systematic Review SN Comprehensive Clinical Medicine 2: 2048-2058. https://doi.org/10.1007/s42399-020-00521-8.
BioNTech (2020). A Phase 1/2/3, Placebo-Controlled, Randomized, Observer-Blind, Dose-Finding Study to Evaluate the Safety, Tolerability, Immunogenicity, and Efficacy of Sars-CoV-2 RNA Vaccine Candidates against COVID-19 in Healthy Individuals. PF-07302048 (BNT162 RNA-Based COVID-19 Vaccines) Protocol C4591001. November. https://media.tghn.org/medialibrary/2020/11/C4591001_Clinical_Protocol_Nov2020_Pfizer_BioNTech.pdf.
Blumenthal, K. G., Robinson, L. B., Camargo, C. Jr., Shenoy, E. S., Banerji, A., Landman, A. B., Wickner, P. (2021) Acute Allergic Reactions to mRNA COVID-19 Vaccines. Journal of the American Medical Association 325(15):1562-1565. https://doi.org/10.1001/jama.2021.3976.
Bonsell, D. (2021, January 10). Largest Multi-Site Distribution Complex in Defense Department Delivers for Operation Warp. Defense Logistics Agency. Retrieved January 27, 2021, from https://www.dla.mil/AboutDLA/News/NewsArticleView/Article/2467282/largest-warehouse-in-defense-department-delivers-for-operation-warp-speed/Bouabe, H. & Okkenhaug, K. (2013). Gene Targeting in Mice: a Review. Methods in Molecular Biology 2013; 1064: 315-336. https://doi.org/10.1007/978-1-62703-601-6_23.
Brown, R. B. (2021) Outcome Reporting Bias in COVID-19 mRNA Vaccine Clinical Trials.Medicina (Kaunas) 57(3): 199. https://www.doi.org/10.3390/medicina57030199.
Buonsenso, D., Riitano, F., & Valentini, P. (2020). Pediatric Inflammatory Multisystem Syndrome Temporally Related with SARS-CoV-2: Immunological Similarities with Acute Rheumatic Fever and Toxic Shock Syndrome.Frontiers in Pediatrics8: 574. https://doi.org/10.3389/fped.2020.00574.
Bushman, D. M., Kaeser, G. E., Siddoway, B., Westra, J. W., Rivera, R. R., Rehen, S. K. … Chun, J. (2015). Genomic Mosaicism with Increased Amyloid Precursor Protein (APP) Gene Copy Number in Single Neurons from Sporadic Alzheimers Disease Brains. eLife 4: e05116. https://doi.org/10.7554/eLife.05116.
Buzhdygana, T. P., DeOrec, B. J., Baldwin-Leclair, A., Bullock, T. A., McGary, H. M…. Ramirez, S. H. (2020). The SARS-CoV-2 Spike Protein Alters Barrier Function in 2D Static and 3D Microfluidic in-Vitro Models of the Human Blood-Brain Barrier. Neurobiology of Disease 146: 105131. https://doi.org/10.1016/j.nbd.2020.105131.
CDC COVID-19 Response Team; Food and Drug Administration (2021, January 15). Allergic Reactions Including Anaphylaxis After Receipt of the First Dose of Pfizer-BioNTech COVID-19 Vaccine—United States, December 14–23, 2020. Morbidity and Mortality Weekly Report 70(2): 46. https://www.cdc.gov/mmwr/volumes/70/wr/mm7002e1.htm.
CDC COVID-19 Response Team; Food and Drug Administration (2021, January 29). Allergic Reactions Including Anaphylaxis After Receipt of the First Dose of Moderna COVID-19 Vaccine-United States. December 21, 2020 –January 10, 2021. MMWR. Morbidity and Mortality Weekly Report 70(4): 125-129. https://www.cdc.gov/mmwr/volumes/70/wr/mm7004e1.htm.
Campos, J. Slon, L., Mongkolsapaya, J., & Screaton, G. R. (2018). The Immune Response Against Flaviviruses. Nature immunology 19(11): 1189-1198. https://doi.org/10.1038/s41590-018-0210-3.
Carsetti, R., Zaffina, S., Piano Mortari, E., Terreri, S., Corrente, F., Capponi, C., … & Locatelli, F. (2020). Different Innate and Adaptive Immune Responses to SARS-CoV-2 Infection of Asymptomatic, Mild, and Severe Cases.Frontiers in immunology,11, 3365. https://www.frontiersin.org/articles/10.3389/fimmu.2020.610300/full
Carter, M. J., Fish, M., Jennings, A., Doores, K. J., Wellman, P., Seow, J., … Shankar-Hari, M. (2020). Peripheral Immunophenotypes in Children with Multisystem Inflammatory Syndrome Associated with SARS-CoV-2 Infection.Nature Medicine,26(11), 1701-1707. https://doi.org/10.1038/s41591-020-1054-6.
Centers for Disease Control and Prevention. COVID Data Tracker. https://covid.cdc.gov/covid-data-tracker/#vaccinations.Accessed 2/6/21.
Centers for Disease Control and Prevention, Prion Diseases. October 9, 2018. https://www.cdc.gov/prions/index.html.
Centers for Disease Control and Prevention (1990). Vaccine Adverse Events Reporting System [database]. Retrieved February 11, 2021 from https://vaers.hhs.gov/about.html
Chen, W., Yang, B., Li, Z., Wang, P., Chen, Y. & Zhou, H. (2020). Sudden Severe Thrombocytopenia in a Patient in the Recovery Stage of COVID-19. Lancet Haematology 7(8): e624. https://doi.org/10.1016/S2352-3026(20)30175-7.
Cifuentes-Diaz, C., Delaporte, C., Dautréaux,B., Charron, D. & Fardeau, M. (1992) Class II MHC Antigens in Normal Human Skeletal Muscle. Muscle Nerve 15(3): 295-302. https://doi.org/10.1002/mus.880150307.
Classen, J. B. (2021). Review of COVID-19 Vaccines and the Risk of Chronic Adverse Events Including Neurological Degeneration. Journal of Medical-Clinical Research and Reviews 5(4): 1-7. https://foundationforhealthresearch.org/review-of-covid-19-vaccines-and-the-risk-of-chronic-adverse-events/.
Corbett, K. S., Edwards, D.K., Leist, S. R., Abiona, O. M., Boyoglu-Barnum, S., Gillespie, R. A. … Graham, B. S. (2020) SARS-CoV-2 mRNA Vaccine Design Enabled by Prototype Pathogen Preparedness. Nature 586(7830): 567-571. https://doi.org/10.1038/s41586-020-2622-0.
Danielsson, R. & Eriksson, H. (2021, January 7). Aluminium Adjuvants in Vaccines –A Way to Modulate the Immune Response. Seminars in Cell & Developmental Biology. (Epub ahead of print) https://doi.org/10.1016/j.semcdb.2020.12.008.
Decock, M, Stanga, S., Octave, J.-N., Dewachter, I., Smith, S. O., Constantinescu, S. N., and Kienlen-Campard, P. (2016). Glycines from the APP GXXXG/GXXXA Trans-membrane Motifs Promote Formation of Pathogenic A Oligomers in Cells. Frontiers in Aging Neuroscience 8: 107. https://doi.org/10.3389/fnagi.2016.00107.
Dicks, M. D. J., Spencer, A. J., Edwards, N. J., Wadell, G., Bojang, %K., Gilbert, S.C., … Cottingham, M. G. (2012). A Novel Chimpanzee Adenovirus Vector with Low Human Seroprevalence: Improved Systems for Vector Derivation and Comparative Immunogenicity. PLoS ONE 7(7): e40385. https://doi.org/10.1371/journal.pone.0040385.
Doshi, P. (2020). Will COVID-19 Vaccines Save Lives? Current Trials Aren’t Designed to Tell Us. BMJ 371: m4037. https://doi.org/10.1136/bmj.m4037.
Doshi, P. (2021a). Peter Doshi: Pfizer and Moderna’s “95% effective” vaccines—we need more details and the raw data. BMJ blog. Accessed 02/20/2021. https://blogs.bmj.com/bmj/2021/01/04/peter-doshi-pfizer-and-modernas-95-effective-vaccines-we-need-more-details-and-the-raw-data/
Doshi, P. (2021b). Clarification: Pfizer and Moderna’s “95% effective” Vaccines –We Need More Details and the Raw Data. BMJ blog. Accessed 02/20/21. https://blogs.bmj.com/bmj/2021/02/05/clarification-pfizer-and-modernas-95-effective-vaccines-we-need-more-details-and-the-raw-data/
Ehrenfeld, M., Tincani, A., Andreoli, L., Cattalini, M., Greenbaum, A., Kanduc, D. … Shoenfeld, Y. (2020). COVID-19 and Autoimmunity. Autoimmunity Reviews 19(8): 102597. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7289100/
EMA Public Assessment Report on Pfizer-BioNTech Vaccine. (2020). Accessed 5/2/21. https://www.documentcloud.org/documents/20516010-ema-assessment-report-12-21-2020#document/p35/a2023027
Eroshenko, N., Gill, T., Keaveney, M. K., Church, G. M., Trevejo, J. M. & Rajaniemi, H. (2020). Implications of Antibody-dependent Enhancement of Infection for SARS-CoV-2 Countermeasures. Nature Biotechnology 38(7): 789-791. https://doi.org/10.1038/s41587-020-0577-1.
European Medicines Agency. Committee for Medicinal Products for Human Use (CHMP) Assessment report. COVID-19 Vaccine Moderna. Common name: COVID-19 mRNA Vaccine (nucleoside-modified) Procedure. No. EMEA/H/C/005791/0000. March 11 2021. p. 47. https://www.ema.europa.eu/en/documents/assessment-report/covid-19-vaccine-moderna-epar-public-assessment-report_en.pdf
Firdessa-Fite, R. & Creusot, R. J. (2020). Nanoparticles versus Dendritic Cells as Vehicles to Deliver mRNA Encoding Multiple Epitopes for Immunotherapy. Molecular Therapy: Methods & Clinical Development 16: 50-62. https://doi.org/10.1016/j.omtm.2019.10.015.
Franke, C., Ferse, C., Kreye, J., Reincke, S. M., Sanchez-Sendin, E., Rocco, A., … & Pruess, H. (2021). High Frequency of Cerebrospinal Fluid Autoantibodies in COVID-19 Patients with Neurological Symptoms.Brain, Behavior, and Immunity 93: 415-419. https://doi.org/10.1016/j.bbi.2020.12.022.
Fujiwara, Y., Wada, K. & Kabuta, T. (2017). Lysosomal Degradation of Intracellular Nucleic Acids –Multiple Autophagic Pathways. The Journal of Biochemistry 161(2): 145-154. https://doi.org/10.1093/jb/mvw085.
Furer, V., Zisman, D., Kibari, A., Rimar, D., Paran, Y., & Elkayam, O. (2021). Herpes zoster Following BNT162b2 mRNA Covid-19 Vaccination in Patients with Autoimmune Inflammatory Rheumatic Diseases: a Case Series. Rheumatology keab345. April 12 [Epub ahead of print] https://doi.org/10.1093/rheumatology/keab345.
Galeotti, C., & Bayry, J. (2020). Autoimmune and Inflammatory Diseases Following COVID-19.Nature Reviews Rheumatology,16(8), 413-414. https://doi.org/10.1038/s41584-020-0448-7.
Gallie, D.R., (1991) The Cap and Poly(A) Tail Function Synergistically to Regulate mRNA Translational Efficiency. Genes & Development 5: 2108–2116. https://doi.org/10.1101/gad.5.11.2108.
Ganson, N. J., Povsic, T. J., Sullenger, B. A., Alexander, J. H., Zelenkofske, S. L., … Hershfield, M. S. (2016). Pre-existing Anti–Polyethylene Glycol Antibody Linked to First-Exposure Allergic Reactions to Pegnivacogin, A PEGylated RNA Aptamer. Journal of Allergy and ClinicalImmunology 137(5): 1610-1613. https://doi.org/10.1016/j.jaci.2015.10.034.
Garvey, L. H., & Nasser, S. (2020, December 17) Allergic Reactions to the First COVID-19 Vaccine: is Polyethylene Glycol (PEG) the Culprit? British Journal of Anaesthesia. Epub ahead of print. https://doi.org/10.1016/j.bja.2020.12.020.
Gao, Z., Xu, Y., Sun, C., Wang, X., Guo, Y., Qiu, S., & Ma, K. (2020). A systematic review of asymptomatic infections with COVID-19.Journal of Microbiology, Immunology and Infection 54(1): 12-16. https://www.sciencedirect.com/science/article/pii/S1684118220301134.
Gao, Z. W., Zhang, H. Z., Liu, C., & Dong, K. (2021). Autoantibodies in COVID-19: Frequency and Function. Autoimmune Reviews 20(3): 102754. https://doi.org/10.1016/j.autrev.2021.102754.
Geuking, M. B., Weber, J., Dewannieux, M., Gorelik, E., Heidmann, T., Hengartner, H., … Hangartner, L. (2009). Recombination of Retrotransposon and Exogenous RNA Virus Results in Nonretroviral cDNA Integration. Science 323(5912): 393-6. https://doi.org/10.1126/science.1167375.
Goddek, S. (2020). Vitamin D3 and K2 and Their Potential Contribution to Reducing the COVID-19 Mortality Rate. International Journal of Infectious Diseases 99: 286-290. https://doi.org/10.1016/j.ijid.2020.07.080.
Gordon, J. W., Scangos, G. A., Plotkin, D. J., Barbosa, J. A. & Ruddle, F.H. (1980). Genetic Transformation of Mouse Embryos by Microinjection of Purified DNA. Proceedings of the National Academy of Sciences USA.77: 7380-84. https://doi.org/10.1073/pnas.77.12.7380.
Grady, D. & Mazzei, P. (2021). Doctor’s DeathAfter COVID Vaccine Is Being Investigated. New York Times Jan. 12. https://www.nytimes.com/2021/01/12/health/covid-vaccine-death.html.
Grady, D. (2021). A Few Covid Vaccine Recipients Developed a Rare Blood Disorder. New York Times Feb. 8. https://www.nytimes.com/2021/02/08/health/immune-thrombocytopenia-covid-vaccine-blood.html.
Haidere, M. F., Ratan, Z. A., Nowroz, S., Zaman, S. B., Jung, Y. J., Hosseinzadeh, H., & Cho, J. Y. (2021). COVID-19 Vaccine: CriticalQuestions with Complicated Answers. Biomolecules & therapeutics, 29(1), 1. https://doi.org/10.4062/biomolther.2020.178.
Hamad, I., Hunter, A. C., Szebeni, J. & Moghimi, S. M. (2008). Poly (Ethylene Glycol)s Generate Complement Activation Products in Human Serum through Increased Alternative Pathway Turnover and a MASP-2-Dependent Process. Molecular immunology 46(2): 225-232. https://doi.org/10.1016/j.molimm.2008.08.276.
Hawkes, R. A. (1964). Enhancement of the Infectivity of Arboviruses by Specific Antisera Produced in Domestic Fowls. Australian Journal of Experimental Biology and Medical Science 42(4): 465-482. https://doi.org/10.1038/icb.1964.44.
Ho, W., Gao, M.,Li, F., Li, J., Zhang, X.-Q. & Xu, X. (2021, January 18). Next‐Generation Vaccines: Nanoparticle‐Mediated DNA and mRNA Delivery. Advanced Healthcare Materials 10(8): e2001812. https://doi.org/10.1002/adhm.202001812.
Hong, L., Wang, Z., Wei, X., Shi, J. & Li, C. (2020). Antibodies Against Polyethylene Glycol in Human Blood: A Literature Review. Journal of Pharmacological and Toxicological Methods 102: 106678. https://doi.org/10.1016/j.vascn.2020.106678.
Hubert, B. Reverse Engineering the source code of the BioNTech/Pfizer SARS-CoV-2 Vaccine. Dec. 25, 2020. https://berthub.eu/articles/posts/reverse-engineering-source-code-of-the-biontech-pfizer-vaccine/
Idrees D, Kumar V. SARS-CoV-2 spike protein interactions with amyloidogenic proteins: Potential clues to Neurodegeneration. Biochemical and Biophysical Re-search Com-munications. 2021; 554: 94-98. https://www.doi.org/10.1016/j.bbrc.2021.03.100.
Jackson, L. A., Anderson, E. J., Rouphael, N. G., Roberts, P. C., Makhene, M., Coler, R. N. … Beigel, J. H. (2020). An mRNA Vaccine against SARS-CoV-2 Preliminary Report. The New England Journal of Medicine 383: 1920-31. https://doi.org/10.1056/NEJMoa2022483.
Jacobs, J. & Armstrong, D. (2020, April 29) Trump’s `Operation Warp Speed’ Aims to Rush Coronavirus Vaccine Bloomberg.Retreived February 11 from https://www.bloomberg.com/news/articles/2020-04-29/trump-s-operation-warp-speed-aims-to-rush-coronavirus-vaccine.
Jaenisch R. (1976). Germ Line Integration and Mendelian Transmission of the Exogenous Moloney Leukemia Virus. Proceedings of the National Academy of Sciences of the United States of America 73: 1260-1264. https://doi.org/10.1073/pnas.73.4.1260.
Jansen, A. J. G., Spaan, T., Low, H. Z., Di Iorio D., van den Brand, J., Malte Tieke, M., … van der Vries, E. (2020). Influenza-Induced Thrombocytopenia is Dependent on the Subtype and Sialoglycan Receptor and Increases with Virus Pathogenicity. Blood Advances 4(13): 2967-2978. https://doi.org/10.1182/bloodadvances.2020001640.
Jiang, Y., Arase, N., Kohyama, M., Hirayasu,K., Suenaga, T., Jin, H., … Hisashi Arase , H. (2013) Transport of Misfolded Endoplasmic Reticulum Proteins to the Cell Surface by MHC Class II Molecules. International Immunology 25(4): 235-246. https://doi.org/10.1093/intimm/dxs155
Kaeser, G. E. & Chun, J. (2020). Mosaic Somatic Gene Recombination as a Potentially Unifying Hypothesis for Alzheimers Disease. Frontiers in Genetics 11: 390. https://doi.org/10.3389/fgene.2020.00390.
Kakarla, R., Hur, J., Kim, Y. J., Kim, J., and Chwae, Y.-J. (2020). Apoptotic Cell-derived Exosomes: Messages from Dying Cells. Experimental & Molecular Medicine 52: 16 https://www.doi.org/10.1038/s12276-019-0362-8.
Karikó, K., Muramatsu, H., Welsh, F. A., Ludwig, J., Kato, H., Akira, S. & Weissman, D. (2008). Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Molecular Therapy 16(11): 1833-1840. https://doi.org/10.1038/mt.2008.200.
Kelso, J. M. (2021) Anaphylactic Reactions to Novel mRNA SARS-CoV-2/COVID-19 Vaccines. Vaccine 39(6): 865–867. https://doi.org/10.1016/j.vaccine.2020.12.084.
Kemp, S. A., Collier, D. A. Datir, R. P., Ferreira, I. A. T. M. Gayed, S., Jahun, A. … Gupta, R. K. (2021) SARS-CoV-2 Evolution during Treatment of Chronic Infection. Nature 2021 Apr;592(7853):277-282. https://doi.org/10.1038/s41586-021-03291-y.
Khodakaram-Tafti, A. & Farjanikish, G. H. (2017) Persistent Bovine Viral Diarrhea Virus (BVDV) Infection in Cattle Herds. Iranian Journal of Veterinary Research, Shiraz University 18(3): 154-163. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5674437/.
Kosuri, S. & Church, G. M., Large-Scale de Novo DNA Synthesis: Technologies and Applications. Nature Methods 2014; 11 (5): 499–507. https://doi.org/10.1038/nmeth.2918.
Koupenova, M., Corkrey, H. A., Vitseva, O., Manni, G., Pang, C. J., Clancy, L. … Freedman, J. E. (2019). The Role of Platelets in Mediating a Response to Human Influenza Infection. Nature Communications 2019;10: 1780. https://doi.org/10.1038/s41467-019-09607-x.
Kozma, G. T., Mészáros, T., Vashegyi, I., Fülöp, T., Örfi, E., Dézsi, L., … Szebeni, J. (2019). Pseudo-anaphylaxis to Polyethylene Glycol (PEG)-Coated Liposomes: Roles of Anti-PEG IgM and Complement Activation in a Porcine Modelof Human Infusion Reactions. ACS Nano 13(8): 9315-9324. https://doi.org/10.1021/acsnano.9b03942.
Ku, C.-C., Chang, Y.-H., Chien, Y., & Lee, T.-L. (2016). Type I Interferon Inhibits Varicella-zoster Virus Replication by Interfering with the Dynamic Interaction between Mediator and IE62 within Replication Compartments. Cell & Bioscience 6: 21. https://doi.org/10.1186/s13578-016-0086-6.
Kuba, K., Imai, Y., Rao, S., Gao, H., Guo, F., Guan, B. … Penninger, J. M. (2005). A Crucial Role of Angiotensin Converting Enzyme 2 (ACE2) in SARS Coronavirus-Induced Lung Injury. Natural Medicine 11: 875-879. https://doi.org/10.1038/nm1267.
Kudla, G., Lipinski, L., Caffin, F., Helwak, A., Zylicz, M. (2006) High Guanine and Cytosine Content Increases mRNA Levels in Mammalian Cells. PlOS Biology 4(6): e180. https://doi.org/10.1371/journal.pbio.0040180.
Kullaya, V., de Jonge, M. E., Langereis, J. D., van der Gaast-de Jongh, C. E., Büll, C., Adema, G. J. … van der Ven, A. J. (2018). Desialylation of Platelets by Pneumococcal Neuraminidase A Induces ADP-Dependent Platelet Hyperreactivity. Infection and Immunity 86(10): e00213-18. https://doi.org/10.1128/IAI.00213-18.
Lambrecht, B. N., Kool, M., Willart, M. A. M. & Hammad, H. (2009). Mechanism of Action of Clinically Approved Adjuvants. Current opinion in immunology 21.1 (2009): 23-29. https://doi.org/10.1016/j.coi.2009.01.004.
Lazzaro, S., Giovani, C., Mangiavacchi, S., Magini,D., Maione, D., Baudner, B., … Buonsanti, C. (2015). CD8 T-cell Priming upon mRNA Vaccination is Restricted to Bone-marrow-derived Antigen-presenting Cells and May Involve Antigen Transfer from Myocytes.Immunology 146: 312-326. https://doi.org/10.1111/imm.12505.
Michael Klompas, Steve Bernstein, and Harvard Pilgrim Health Care, Inc. 2010. “Electronic Support for Public Health–Vaccine Adverse Event Reporting System (ESP-VAERS).” Rockville, MD: Harvard Pilgrim Health Care, Inc. https://healthit.ahrq.gov/sites/default/files/docs/publication/r18hs017045-lazarus-final-report-2011.pdf.
Lederer, K., Castaño, D., Gómez Atria, D., Oguin T. H., III, Wang, S., Manzoni, T. B., … (2020).SARS-CoV-2 mRNA Vaccines Foster Potent Antigen-Specific Germinal Center Responses Associated with Neutralizing Antibody Generation.Immunity 53: 1281-1295. https://doi.org/10.1016/j.immuni.2020.11.009.
Lee, S. H., Cha, J. M., Lee, J. I., Joo, K. R., Shin, H. P., Baek, I. H. … Cho, J. L. (2015). Anaphylactic Shock Caused by Ingestion of Polyethylene Glycol. Intestinal research 13(1): 90-94. https://doi.org/10.5217/ir.2015.13.1.90.
Lee, W. S., Wheatley, A. K., Kent, S. J. & DeKosky, B. J. (2020). Antibody-Dependent Enhancement and SARS-CoV-2 Vaccines and Therapies. Nature Microbiology 5(10): 1185-1191. https://doi.org/10.1038/s41564-020-00789-5.
Lema Tomé, C. M., Tyson, T., Rey, N. L., Grathwohl, S., Britschgi, M. and Brundin, P. (2013). Inflammation and α-Synuclein Prion-like Behavior in Parkinson’s Disease –Is There a Link? Molecular Neurobiology 47: 561-574. https://www.doi.org/10.1007/s12035-012-8267-8.
Lesbats, P., Engelman, A. N. & Cherepanov, P. (2016). Retroviral DNA Integration. Chemical Reviews 2016 116(20): 12730012757. https://doi.org/10.1021/acs.chemrev.6b00125.
Liang, J., Zhu, H., Wang, X., Jing, B., Li, Z., Xia, X. … Sun, B. (2020). Adjuvants for Coronavirus Vaccines. Frontiers in Immunology 11: 2896. https://doi.org/10.3389/fimmu.2020.589833.
Lila, A. S., Shimizu, A. T. & Ishida, T. (2018). PEGylation and Anti-PEG Antibodies. Engineering of Biomaterials for Drug Delivery Systems. Woodhead Publishing 51-68. https://doi.org/10.1016/B978-0-08-101750-0.00003-9.
Limanaqi, F., Letizia Busceti, C., Biagioni, F., Lazzeri, G., Forte, M., Schiavon, S. … Fornai, F. (2020). Cell Clearing Systems as Targets of Polyphenols in Viral Infections: Potential Implications for COVID-19 Pathogenesis. Antioxidants 9: 1105. https://doi.org/10.3390/antiox9111105.
Lindsay, K. E., Bhosle, S. M., Zurla, C., Beyersdorf, J., Rogers, K. A., Vanover D. & Xiao, P. (2019). Visualization of Early Events in mRNA Vaccine Delivery in Non-Human Primates via PET–CT and Near-Infrared Imaging. Nature Biomedical Engineering 3: 371-380. https://doi.org/10.1038/s41551-019-0378-3.
Lipp, E., von Felten, A., Sax, H., Mller, D. & Berchtold, P. (1998). Antibodies Against Platelet Glycoproteins and Antiphospholipid Antibodies in Autoimmune Thrombocytopenia. European Journal of Haematology 60(5): 283-8. https://doi.org/10.1111/j.1600-0609.1998.tb01041.x.
Liu, L., Wei, Q., Lin, Q., Fang, J., Wang, H., Kwok, H., … Chen, Z. (2019). Anti–spike IgG Causes Severe Acute Lung Injury by Skewing Macrophage Responses During Acute SARS-CoV Infection. JCI Insight 4(4): e123158. https://doi.org/10.1172/jci.insight.123158.
Liu, M. A. (2019). A Comparison of Plasmid DNA and mRNA as Vaccine Technologies. Vaccines (Basel) 7(2): 37. https://doi.org/10.3390/vaccines7020037.
Liu, S., Hossinger, A., Gbbels, S., and Ina M. Vorberga, I. M. (2017). Prions on the Run: How Extracellular Vesicles Serve as Delivery Vehicles for Self-templating Protein Aggregates. Prion 11(2): 98-112. https://www.doi.org/10.1080/19336896.2017.1306162.
Liu, Y., Liu, J., Xia, H., Zhang, X., Fontes-Garfias, C. R., Swanson, K. A. … Shi, P.-Y. (2021). Neutralizing Activity of BNT162b2-Elicited Serum. N Engl J Med 384: 1466-1468. https://doi.org/10.1056/NEJMc2102017.
Louis, N., Evelegh, C., Graham, F. L. (1997) Cloning and Sequencing of the Cellular-Viral Junctions from the Human Adenovirus Type 5 Transformed 293 Cell Line. Virology 233: 423-429. https://doi.org/10.1006/viro.1997.8597.
Lu, J., Lu, G., Tan, S., Xia, J., Xiong, H., Yu, X. … Lin, J. (2020). A COVID-19 mRNA Vaccine Encoding SARS-CoV-2 Virus-like Particles Induces a Strong Antiviral-like Immune Response in Mice. Cell Research 30: 936-939. https://doi.org/10.1038/s41422-020-00392-7.
Lu, L., Li, J., Moussaoui, M. & Boix, E. (2018). Immune Modulation by Human Secreted RNases at the Extracellular Space. Frontiers in Immunology 9: 1012. https://doi.org/10.3389/fimmu.2018.01012.
Lu, L. L., Suscovich, T. J., Fortune, S. M. & Alter G. (2018b). Beyond Binding: Antibody Effector Functions in Infectious Diseases. Nature Reviews Immunology18(1): 46-61. https://doi.org/10.1038/nri.2017.106.
Lucchetti, D., Santini, G., Perelli, L., Ricciardi-Tenore, C., Colella, F., Mores, N., … Montuschi, P. (2021). Detection and Characterization of Extracellular Vesicles in Exhaled Breath Condensate and Sputum of COPD and Severe Asthma Patients. European Respiratory Journal Apr 1; 2003024. [Epub ahead of print]. https://www.doi.org/10.1183/13993003.03024-2020.
Luganini, A. & Gribaudo, G. (2020). Retroviruses of the Human Virobiota: The Recycling of Viral Genes and the Resulting Advantages for Human Hosts During Evolution. Frontiers inMicrobiology 11: 1140. https://doi.org/10.3389/fmicb.2020.01140.
Lyons-Weiler, J. (2020). Pathogenic Priming Likely Contributes to Serious and Critical Illness and Mortality in COVID-19 via Autoimmunity. Journal of Translational Autoimmunity 3: 100051. https://www.sciencedirect.com/science/article/pii/S2589909020300186.
Mahose, E. (2021) Covid-19: Booster Dose will beNeeded in Autumn to Avoid Winter Surge, Says Government Adviser. BMJ 372: n664. https://doi.org/10.1136/bmj.n664.Marino, M., Scuderi, F., Provenzano, C. & Bartoccioni, E. (2011) Skeletal Muscle Cells: fromLocal Inflammatory Response to Active Immunity. Gene Therapy 18: 109-116. https://doi.org/10.1038/gt.2010.124.
Matsuno, H., Yudoh, K., Katayama, R., Nakazawa, F., Uzuki, M., Sawai, T., … Kimura, T. (2002). The Role of TNF-αiin the Pathogenesis of Inflammation and Joint Destruction in Rheumatoid Arthritis (RA): a Study Using a Human RA/SCID Mouse Chimera. Rheumatology (Oxford) 41(3): 329-37. https://doi.org/10.1093/rheumatology/41.3.329.
McClintock, B. (1965). Components of Action of the Regulators Spm and Ac. Carnegie Institution of Washington Year Book 64: 527-536. http://repository.cshl.edu/id/eprint/34634/.
McNeil, M. M., Weintraub, E. S., Duffy, J., Sukumaran, L., Jacobsen, S. J., Klein, N. P. … DeStefano, F. (2016). Risk of Anaphylaxis after Vaccination in Children and Adults. The Journal of Allergy and Clinical Immunology 137(3): 868-78. https://doi.org/10.1016/j.jaci.2015.07.048.
Mehta, N., Sales, R. M., Babagbemi, K., Levy, A. D., McGrath, A. L., Drotman, M. & Dodelzon. K. (2021). Unilateral axillary Adenopathy in the setting of COVID-19 vaccine. Clinical Imaging 75: 12-15. https://doi.org/10.1016/j.clinimag.2021.01.016.
Mi, S., Lee, X., Li, X., Veldman, G. M., Finnerty, H., Racie, L. … McCoy, J. M. (2000). Syncytin is a Captive Retroviral Envelope Protein Involved in Human Placental Morphogenesis. Nature 403(6771): 785-9. https://doi.org/10.1038/35001608.
Moderna. mRNA Platform: Enabling Drug Discovery & Development. 2020. https://www.modernatx.com/mrna-technology/mrna-platform-enabling-drug-discovery-development
Mohamed, M., Lila, A. S., Shimizu, T., Alaaeldin, E., Hussein, A., Sarhan, H. A., Szebeni, J. & Ishida, T. (2019).PEGylated Liposomes: Immunological Responses. Science and Technology of Advanced Materials 20(1): 710-724. https://doi.org/10.1080/14686996.2019.1627174.
Morens, D. M. (1994). Antibody-dependent Enhancement of Infection and the Pathogenesis of Viral Disease. Clinical Infectious Diseases 19(3): 500-512, https://doi.org/10.1093/clinids/19.3.500.
Mueller, B. K., Subramaniam, S., and Senes, A. (2014). A Frequent, GxxxG-mediated, Transmembrane Association Motif is Optimized for the Formation of Interhelical Cα-H Hydrogen Bonds. PNAS E888-E895. Proceedings of the Natural Academy of Sciences USA 111(10): E888-95. https://doi.org/10.1073/pnas.1319944111.
National Institutes of Health (December 11, 2020). NIH-Moderna COVID-19 Vaccine Shows Promising Interim. Results. NIH Record Vol. LXXII, No. 25. Retrieved January 27, 2021 from https://nihrecord.nih.gov/2020/12/11/nih-moderna-covid-19-vaccine-shows-promising-interim-results
Navarra, A., Albani, E., Castellano, S., Arruzzolo L., & Levi-Setti P. E. (2020). Coronavirus Disease-19 Infection: Implications on Male Fertility and Reproduction. Frontiers in Physiology 11: 574761. https://www.doi.org/10.3389/fphys.2020.574761.
Ndeupen, S., Qin, Z., Jacobsen, S., Estanbouli, H., Bouteau, A., & Igyártó, B.Z. (2021) The mRNA-LNP Platform’s Lipid Nanoparticle Component Used in Preclinical Vaccine Studies is Highly Inflammatory. bioRxiv 2021.03.04.430128. https://doi.org/10.1101/2021.03.04.430128.
Norling, K., Bernasconi, V., Hernández, V. A., Parveen, N., Edwards, K., Lycke, N. Y. … Bally. M. (2019). Gel Phase 1,2-Distearoyl-sn-glycero-3-phosphocholine-Based Liposomes Are Superior to Fluid Phase Liposomes at Augmenting Both Antigen Presentation on Major Histocompatibility Complex Class II and Costimulatory Molecule Display by Dendritic Cells in Vitro. ACS Infectious Diseases 5(11): 1867-1878. https://doi.org/10.1021/acsinfecdis.9b00189.
Oller, J. W., Jr. (2010). The Antithesis of Entropy: Biosemiotic Communication from Genetics to Human Language with Special Emphasis on the Immune Systems. Entropy 12: 631-705. https://www.doi.org/10.3390/e12040631.
Palucka, A. K., Blanck, J. P., Bennett, L., Pascual, V,, Banchereau, J. (2005) Cross-regulation of TNF and IFN-αin Autoimmune Diseases. Proceedings of the National Academy of Sciences USA 102: 3372-3377. https://doi.org/10.1073/pnas.0408506102.
Pellionisz, A. J. (2012). The Decade of Fractogene: From Discovery to Utility -Proofs of Concept Open Genome-Based Clinical Applications. International Journal of Systemics, Cybernetics and Informatics 12-02: 17-28. http://www.junkdna.com/pellionisz_decade_of_fractogene.pdf.
Peron, J. P. S. & Nakaya, H. (2020). Susceptibility of theElderly to SARS-CoV-2 Infection: ACE-2 Overexpression, Shedding, and Antibody-dependent Enhancement (ADE). Clinics (Sao Paulo) 75: e1912. https://doi.org/10.6061/clinics/2020/e1912.
Pittoggi, C., Beraldi, R., Sciamanna, I., Barberi, L., Giordano, R., Magnano, A. R.& Spadafora C (2006). Generation of Biologically Active Retro-genes upon Interaction of Mouse Spermatozoa with Exogenous DNA. Molecular Reproduction and Development 73(10): 1239-46. https://doi.org/10.1002/mrd.20550.
Povsic, T. J., Lawrence, M. G., Lincoff, A. M., Mehran, R., Rusconi, C. P. … REGULATE-PCI Investigators. (2016). Pre-existing Anti-PEG Antibodies are Associated with Severe Immediate Allergic Reactions to Pegnivacogin, a PEGylated Aptamer. Journal of Allergy and Clinical Immunology 138(6): 1712-1715. https://doi.org/10.1016/j.jaci.2016.04.058.
Pray, L. (2008) Transposons, or Jumping Genes: Not Junk DNA? Nature Education 1(1): 32. https://www.nature.com/scitable/topicpage/transposons-or-jumping-genes-not-junk-dna-1211/.Prusiner, S. B. (1982). Novel proteinaceous infectious particles cause scrapie Science 216(4542): 136-44. https://www.doi.org/10.1126/science.6801762.
Puga, I., Cols, M., Barra, C. M., He, B.,Cassis, L., Gentile, M. … Cerutti, A. (2011). B Cell-helper Neutrophils Stimulate the Diversification and Production of Immunoglobulin in the Marginal Zone of the Spleen. Natural Immunology 13(2): 170-80. https://doi.org/10.1038/ni.2194.
Pushparajah, D., Jimeneza, S., Wong, S., Alattas, H., Nafissi, N. & Slavcev, R. A. (2021) Advances in Gene-Based Vaccine Platforms to Address the COVID-19 Pandemic. Advanced Drug Delivery Reviews 170: 113-141. https://doi.org/10.1016/j.addr.2021.01.003.
Rico-Campà, A., Martínez-González, M. A., Alvarez-Alvarez, I., de Deus Mendonça, R., de la Fuente-Arrillaga, C., Gómez-Donoso, C. & Bes-Rastrollo, M. (2019). Association Between Consumption of Ultra-Processed Foods and All Cause Mortality: SUN Prospective Cohort Study. Journal of Infection and Public Health 13(10): 1373-1380. https://pubmed.ncbi.nlm.nih.gov/31142450/
Rocha, E. P. C., & Danchin, A. (2002). Base composition bias might result from competition for metabolic resources. Trends in Genetics, 18(6), 291–294. https://doi.org/10.1016/S0168-9525(02)02690-2
Sarohan, A. R. (2020). COVID-19: Endogenous Retinoic Acid Theory and Retinoic Acid Depletion Syndrome. Medical Hypotheses 144: 110250. https://www.doi.org/10.1016/j.mehy.2020.110250.
Schiaffino, M. T., Di Natale, M., García-Martínez, E., Navarro, J., Muñoz-Blanco, J. L., Demelo-Rodríguez, P., & Sánchez-Mateos, P. (2020). Immunoserologic Detection and Diagnostic Relevance of Cross-reactive Autoantibodies in Coronavirus Disease 2019 Patients.The Journal of Infectious Diseases,222(9), 1439-1443. https://doi.org/10.1093/infdis/jiaa485.
Schlake, T., Thess, A., Fotin-Mleczek, M. & Kallen, K.-J. (2012). Developing mRNA-vaccine technologies, RNA Biology 9: 1319–1330. https://doi.org/10.4161/rna.22269.
Sellaturay, P., Nasser, S., & Ewan, P. (2020). Polyethylene Glycol (PEG)-Induced Anaphylactic Reaction During Bowel Preparation. ACG Case Reports Journal 2(4) 216-217. https://doi.org/10.14309/crj.2015.63.
Sellaturay, P., Nasser, S., & Ewan, P. (2020). Polyethylene Glycol–Induced Systemic Allergic Reactions (Anaphylaxis). The Journal of Allergy and Clinical Immunology: In Practice 9(2): 670-675. https://doi.org/10.1016/j.jaip.2020.09.029.
Shaw, C.A. (2021). The Age of COVID-19: Fear, Loathing, and the New Normal. International Journal of Vaccine Theory, Practice, and Research 1: 98-142. https://ijvtpr.com/index.php/IJVTPR/article/view/11.
Shukla, R., Ramasamy, V., Shanmugam, R. K., Ahuja, R. & Khanna, N. (2020). Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Frontiers in Cellular and Infection Microbiology 10: 572681. https://doi.org/10.3389/fcimb.2020.572681.
Slimani, Y., Abbassi, R., El Fatoiki, F. Z., Barrou, L., & Chiheb, S. (2021). Systemic Lupus Erythematosus and Varicella‐Like Rash Following COVID‐19 in a Previously Healthy Patient.Journal of Medical Virology93(2): 1184-1187. https://doi.org/10.1002/jmv.26513.
Steele, E. J., Gorczynski, R. M., Lindley, R. A., Liu, Y., Temple, R., Tokoro, G., … Wickramasinghe, , N. C. (2019). Lamarck and Panspermia -On the Efficient Spread of Living Systems Throughout the Cosmos. Progress in Biophysics and Molecular Biology 149: 10-32. https://doi.org/10.1016/j.pbiomolbio.2019.08.010.
Steiner, J. A., Angot, E., and Brundin, P. (2011). A Deadly Spread: Cellular Mechanisms of α-Synuclein Transfer. Cell Death and Differentiation 18: 1425-1433. https://www.doi.org/10.1038/cdd.2011.53.
Stokes, A., Pion, J., Binazon, O., Laffont, B., Bigras, M., Dubois, G. … Rodriguez L.-A. (2020). Nonclinical Safety Assessment of Repeated Administration and Biodistribution of a Novel Rabies Self-amplifying mRNA Vaccine in Rats. Regulatory Toxicology and Pharmacology 113: 104648. https://doi.org/10.1016/j.yrtph.2020.104648.
Su, J. R., Moro, P. L., Ng, C. S., Lewis, P. W., Said, M. A., & Cano, M.V. (2019). Anaphylaxis after vaccination reported to theVaccine Adverse Event Reporting System, 1990-2016.Journal of Allergy and Clinical Immunology 143(4): 1465-1473. https://doi.org/10.1016/j.jaci.2018.12.1003.
Sun, R.-J. & Shan, N.-N. (2019). Megakaryocytic Dysfunction in Immune Thrombocytopenia is Linked to Autophagy Cancer Cell International 19: 59. https://doi.org/10.1186/s12935-019-0779-0.
Suzuki, Y. J. & Gychka, S. G. (2021). SARS-CoV-2 Spike Protein Elicits Cell Signaling in Human Host Cells: Implications for Possible Consequences of COVID-19 Vaccines. Vaccines 9: 36. https://doi.org/10.3390/vaccines9010036.
Suzuki, Y. J. (2020). The Viral Protein Fragment Theory of COVID-19 Pathogenesis. Medical Hypotheses 144: 110267. https://doi.org/10.1016/j.mehy.2020.110267.
Suzuki, Y. J., Nikolaienko, S. I., Dibrova, V. A., Dibrova, Y. V., Vasylyk, V. M., Novikov, M. Y. … Gychka, S. G. (2021). SARS-CoV-2 Spike Protein-Mediated Cell Signaling in Lung Vascular Cells. Vascular Pharmacology 137: 106823. https://doi.org/10.1016/j.vph.2020.106823.
Suzuki, Y.J., Nikolaienko, S.I., Dibrova, V.A., Dibrova, Y.V., Vasylyk, V.M., Novikov, M.Y. … Gychka, S.G. (2020). SARS-CoV-2 Spike Protein-Mediated Cell Signaling in Lung Vascular Cells. Vascular Pharmacology 137: 106823. https://www.doi.org/10.1016/j.vph.2020.106823.
Takada, A., Feldmann, H., Ksiazek, T. G. & Kawaoka, Y. (2003). Antibody-Dependent Enhancement of Ebola Virus Infection. Virology 77(13): 7539-7544. https://doi.org/10.1128/JVI.77.13.7539-7544.2003.
Temin, H. M. and Mizutani, S. (1970). RNA-dependent DNA polymerasein virions of Rous Sarcoma Virus. Nature 226: 1211–3. https://www.doi.org/10.1038/2261211a0.
Tetz, G. and Tetz, V. (2020). SARS-CoV-2 Prion-Like Domains in Spike Proteins Enable Higher Affinity to ACE2. Preprints 2020030422. https://www.doi.org/10.20944/preprints202003.0422.v1.
Tetz, G. and Tetz,V (2018). Prion-like Domains in Eukaryotic Viruses. Scientific Reports 8: 8931. https://doi.org/10.1038/s41598-018-27256-w.
U.S. Department of Health and Human Services, Food and Drug Administration. Center for Biologics Evaluation and Research. (2020, June) Development and Licensure of Vaccines to Prevent COVID-19 Guidance for Industry. Retrieved February 11, 2021 from https://www.fda.gov/regulatory-information/search-fda-guidance-documents/development-and-licensure-vaccines-prevent-covid-19.
US Food and Drug Administration (2021). Pfizer-BioNTech COVID-19 Vaccine EUA Fact Sheet for Healthcare Providers Administering Vaccine (Vaccination Providers). https://www.fda.gov/media/144413
Verma, S., Saksena, S. & Sadri-Ardekani, H. (2020). ACE2 Receptor Expression in Testes: Implications in Coronavirus Disease 2019 Pathogenesis. Biology of Reproduction 103(3): 449-451. https://doi.org/10.1093/biolre/ioaa080
Vlachoyiannopoulos, P. G., Magira, E., Alexopoulos, H., Jahaj, E., Theophilopoulou, K., Kotanidou, A., & Tzioufas, A. G. (2020). Autoantibodies Related to Systemic Autoimmune Rheumatic Diseases in Severely IllPatients with COVID-19.Annals of the Rheumatic Diseases79(12): 1661-1663. http://dx.doi.org/10.1136/annrheumdis-2020-218009.
Vaidya, M. and Sugaya, K (2020). DNA Associated with Circulating Exosomes as a Biomarker for Glioma. Genes 11: 1276. https://www.doi.org/10.3390/genes11111276.
Vojdani, A., & Kharrazian, D. (2020). Potential Antigenic Cross-Reactivity Between SARS-CoV-2 and Human Tissue with a Possible Link to an Increase in Autoimmune Diseases.Clinical Immunology (Orlando, Fla.)217: 108480. https://doi.org/10.1016/j.clim.2020.108480.
Vojdani, A., Vojdani, E., & Kharrazian, D. (2021). Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins with Tissue Antigens: Implications for Autoimmune Diseases.Frontiers in Immunology11: 3679. https://doi.org/10.3389/fimmu.2020.617089.
Wadhwa, A., Aljabbari, A., Lokras, A., Foged, C. & Thakur, A. (2020). Opportunities and Challenges in the Delivery of mRNA-based Vaccines. Pharmaceutics 12(2): 102. https://doi.org/10.3390/pharmaceutics12020102.
Wallukat, G., Hohberger, B., Wenzel, K.,Fürst, J.,Schulze-Rothe, S., Wallukat, A. … Müller, J. (2021). Functional Autoantibodies against G-protein CoupledReceptors in Patients with Persistent Post-COVID-19 Symptoms. Journal of Translational Autoimmunity 4: 100100. https://doi.org/10.1016/j.jtauto.2021.100100.
Walter, U., Tsiberidou, P., Kersten, M., Storch, A., and Lohle, M. (2018). Atrophy of the Vagus Nerve in Parkinsons Disease Revealed by High-resolution Ultrasonography. Frontiers in Neurology 9:805. https://www.doi.org/10.3389/fneur.2018.00805.
Wan, Y., Shang, J., Sun, S., Tai, W., Chen, J., Geng, Q., … & Li, F. (2020). Molecular Mechanism for Antibody-Dependent Enhancement of Coronavirus Entry. Journal of virology, 94(5). https://doi.org/10.1128/JVI.02015-19.
Wang, C.-Y., Ma, S., Bi, S.-J., Su,L., Huang, S.-Y. … Peng, J. (2019). Enhancing Autophagy Protects Platelets in Immune Thrombocytopenia Patients. Ann Transl Med 7(7): 134. https://doi.org/10.21037/atm.2019.03.04.
Wang, Z., Troilo, P. J., Wang, X., Griffiths, T.G. II, Pacchione, S. J., Barnum, A. B., … Ledwith, B. J. (2004). Detectionof integration of plasmid DNA into host genomic DNA following intramuscular injection and electroporation. Gene Therapy 11: 711-721. https://doi.org/10.1038/sj.gt.3302213.
Wang, Z.& Xu, X. (2020). ScRNA-seq Profiling of Human Testes Reveals the Presence of the ACE2 Receptor, a Targetfor SARS-CoV-2 Infection in Spermatogonia, Leydig and Sertoli Cells. Cells 9: 920. https://doi.org/10.3390/cells9040920.
Weickenmeier, J., Jucker, M., Goriely, A., and Kuhl, E. (2019). A Physics-based Model Explains the Prion-like Features of Neurodegeneration in Alzheimer’s Disease, Parkinson’s Disease, and Amyotrophic Lateral Sclerosis. Journal of the Mechanics and Physics of Solids 124: 264-281. https://doi.org/10.1016/j.jmps.2018.10.013.
Weiner, A. M. (2002). SINEs and LINEs: the Art of Biting the Hand that Feeds You. Current Opinions in Cell Biology 14(3): 343-50. https://doi.org/10.1016/s0955-0674(02)00338-1.Wikipedia contributors. (2021, February 13). ELISA. Retrieved February 16, 2021, from Wikipedia, The Free Encyclopedia. https://en.wikipedia.org/w/index.php?title=ELISA&oldid=1006455262.
World Health Organization (2021, January 19). mRNA-1273 Vaccine (Moderna) Against COVID-19 Background Document: Draft Prepared by the Strategic Advisory Group of Experts (SAGE) on Immunization Working Group on COVID-19 vaccines. No. WHO/2019-nCoV/vaccines/mRNA-1273/2021.1. https://policycommons.net/artifacts/1424630/mrna-1273-vaccine-moderna-against-covid-19-background-document/
World Health Organization. (2021, January 14). Background document on mRNA vaccine BNT162b2 (Pfizer-BioNTech) against COVID-19. License: CC BY-NC-SA 3.0 IGO. https://apps.who.int/iris/handle/10665/338671.
Wrapp, D., Wang, N., Corbett, K. S., Goldsmith, J. A., Hsieh, C.-L., Abiona, O. … Graham, B. S. (2020). Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science 2020; 367: 1260-3. https://doi.org/10.1126/science.abb2507.
Wu, F., Yan, R., Liu, M., Liu, Z., Wang, Y., Luan, D., … Huang, J. (2020). Antibody-Dependent Enhancement (ADE) of SARS-CoV-2 Infection in Recovered COVID-19 Patients: Studies Based on Cellular and Structural Biology Analysis. medRxiv preprint. https://doi.org/10.1101/2020.10.08.20209114.
Wylon, K. Sabine Dölle, S., & Margitta Worm, M. (2016). Polyethylene Glycol as a Cause of Anaphylaxis. Allergy, Asthma & Clinical Immunology 12(1): 1-3. https://doi.org/10.1186/s13223-016-0172-7.
Xu, S., Yang, K., Li, R. & Zhang, L. (2020) mRNA Vaccine Era –Mechanisms, Drug Platform and Clinical Prospection. International Journal of Molecular Science 21(18): 6582. https://doi.org/10.3390/ijms21186582.
Yang, Q. & Lai, S. K. (2015). Anti‐PEG Immunity: Emergence, Characteristics, and Unaddressed Questions. Wiley Interdisciplinary Reviews: Nanomedicine and Nanobiotechnology 7(5): 655-677. https://doi.org/10.1002/wnan.1339.
Young, R., Bekele, T., Gunn, A., Chapman, N., Chowdhary, V., Corrigan, K., … Yamey, G. (2018). Developing New Health Technologies for Neglected Diseases: A Pipeline Portfolio Review and Cost Model. Gates Open Res 2:23. https://doi.org/10.12688/gatesopenres.12817.2.
Zaman, M. (2021). COVID Vaccine Booster Shots Are Coming —Here’s What to Know. https://www.msn.com/en-us/health/medical/covid-vaccine-booster-shots-are-coming-here-s-what-to-know/ar-BB1foY4s. Accessed 5/1/2021.
Zamani, B., Moeini Taba, S.-M. & Shayestehpour, M. (2021). Systemic Lupus Erythematosus Manifestation Following COVID-19: A Case Report. Journal of Medical Case Reports15(1): 1-4. https://doi.org/10.1186/s13256-020-02582-8.
Zeng, C., Zhang, C, Walker, P. G. & Dong, Y. (2020). Formulation and Delivery Technologies for mRNA Vaccines. Current Topics in Microbiology and Immunology June 2. [Epub ahead of print]. https://doi.org/10.1007/82_2020_217.
Zhang, L., Richards, A., Barrasa, M, I., Hughes, S. H., Young, R. A. & Jaenisch, R. (2021). Reverse-transcribed SARS-CoV-2 RNA can Integrate into the Genome of Cultured Human Cells and can be Expressed in Patient-derived Tissues. Proceedings of the National Academy of Sciences 118(21): e2105968118. https://doi.org/10.1073/pnas.2105968118.
Zhang, X. W. & Yap, Y. L. (2004). The 3D Structure Analysis of SARS-CoV S1 Protein Reveals a Link to Influenza Virus Neuraminidase and Implications for Drug and Antibody Discovery. Theochemistry 681(1): 137-141. https://doi.org/10.1016/j.theochem.2004.04.065.
Zhou, Z.-H., Stone, C. A., Jr., Jakubovic, B., Phillips, E. J., Sussman, G., Park, J.-M. … Kozlowski, S. (2020). Anti-PEG IgE in Anaphylaxis Associated with Polyethylene Glycol. The Journal of Allergy and Clinical Immunology in Practice ;9(4): 1731-1733.e3. https://doi.org/10.1016/j.jaip.2020.11.011.
Zimmer, C., Corum, J., Wee, S.-L. Coronavirus Vaccine Tracker. New York Times. Updated Jan. 28, 2021. https://www.nytimes.com/interactive/2020/science/coronavirus-vaccine-tracker.html.
Zuo, Y., Estes, S. K., Ali, R. A., Gandhi, A. A., Yalavarthi, S., Shi, H., … & Knight, J. S. (2020). Prothrombotic Autoantibodies in Serum from Patients Hospitalized with COVID-19. Science Translational Medicine, 12(570): eabd3876. https://doi.org/10.1126/scitranslmed.abd3876.
Legal Disclaimer
The information on the website and in the IJVTPRis not intended as a diagnosis, recommended treatment, prevention, or cure for any human condition or medical procedure that may be referred to in any way.
Users and readers who may be parents, guardians, caregivers, clinicians, or relatives of persons impacted by any of the morbid conditions, procedures, or protocols that may be referred to, must use their own judgment concerning specific applications.
The contributing authors, editors, and persons associated in any capacity with the website and/or with the journal disclaim anyliability or responsibility to any person or entity for any harm, financial loss, physical injury, or other penalty that may stem from any use or application in any context of information, conclusions, research findings, opinions, errors, or any statements found on the website or in the IJVTPR.
The material presented is freely offered to all users who may take an interest in examining it, but how they may choose to apply any part of it, is the sole responsibility of the viewer/user.
If material is quoted or reprinted, users are asked to give credit to the source/author and to conform to the non-commercial, no derivatives, requirements of the Creative Commons License 4.0 NC ND.
Most Covid patients at Israeli hospital fully vaccinated? What does this mean for Australia?
The Spike Protein is the Reason Why the Vaccines Might Kill You